Префронтальна кора
Префронта́льна кора́ (Cortex praefrontalis) — відділ кори великих півкуль головного мозку, є передньою частиною лобових часток і охоплює 8, 9, 10, 11, 12, 13, 14, 24, 25, 32, 44, 45, 46 і 47 поля Бродмана.[2]
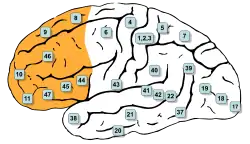 Поля Бродмана 8, 9, 10, 11, 12, 13, 14, 24, 25, 32, 44, 45, 46 і 47 знаходяться в префронтальній корі | |
| Деталі | |
|---|---|
| Артерія | Передня й середня мозкові артерії |
| Вена | Верхній сагітальний синус |
| Ідентифікатори | |
| Латина | Cortex praefrontalis |
| MeSH | D017397 |
| NeuroNames | 2429 |
| NeuroLex ID | nlx_anat_090801 |
| FMA | 224850[1] |
| Анатомічна термінологія | |
Багато авторів знаходили зв'язок між бажанням людини жити, особистістю й функціями префронтальної кори[3]. Ця частина мозку забезпечує планування складної когнітивної поведінки, прояви особистості, прийняття рішень і регулювання соціальної поведінки[4]. Основним призначенням вважається узгодження думок і дій відповідно до внутрішніх цілей[5].
Типовим психологічним терміном для функцій, які виконуються префронтальною корою головного мозку, є виконавча функція. Вона пов'язана зі здібностями виявляти суперечливі думки, прогнозувати майбутні наслідки поточних дій (хороші й погані, хороші та кращі, однакові й різні), діяти в напрямку досягнення певної мети, прогнозувати результати, очікування, засновані на діях, і соціального «контролю» (здатність пригнічувати думки, які можуть привести до соціально неприйнятних результатів).
Лобова кора підтримує засвоєння конкретних правил. Передні частини вздовж ростро-каудальної осі лобової кори підтримують навчання за більш загальними правилами на вищих рівнях абстракції[6].
Визначення
Існують три можливі визначення префронтальної кори:
- гранулярна лобова кора (тобто та, в якій переважають нейрони II й IV шарів — зовнішній і внутрішній гранулярний шар, відповідно — кори великих півкуль)
- зона проєкції медіодорсальних ядер таламуса
- частина лобових часток, електрична стимуляція якої не викликає рухів.
Перше визначення характерне тільки для приматів, бо у неприматів немає IV гранулярного коркового шару; тобто, хоча воно й можна застосувати до людини, але цей термін не буде універсальним[7].
Друге визначення виправдили наступними дослідженнями: виявилося, що медіодорсальні ядра таламуса мають проєкції не тільки в гранулярну лобову кору[8]. Таким чином, визначення скоригували й тепер воно може звучати так: «префронтальна кора — це частина кори великих півкуль, що має сильніші реципрокні зв'язки з медіодорсальними ядрами таламуса, ніж з будь-якими іншими таламічними ядрами».
Третє визначення також не є точним: зони лобових часток, що не відповідають на електростимуляцію руховою активністю, містять і гранулярну, і агранулярну (премоторну) кору[7].
Склад
У наведеній нижче таблиці показані різні способи поділу префронтальної кори на зони Бродмана[2].
| каудальна | ||||||||||||||
Медіальна префронтальна кора головного мозку (mPFC) складається з гранулярних кортикальних областей (медіальні частини полів 9 і 10) і агранулярних областей (поля 24, 25 і 32), які охоплюють передню поясну звивину (поле 24), інфралімбічна кору (поле 25) та прелімбічну кору головного мозку (поле 32)[2].
- Орбітофронтальна кора складається з гранулярних областей (поле 11) і змішаних гранулярних та агранулярних областей (поля 13 і 14)[2].
- Вентролатеральна префронтальна кора складається з полів 12, 44, 45 і 47.
- Дорсолатеральна префронтальна кора складається з латеральної частини поля 9 і всього поля 46.
- Каудальна префронтальна кора складається з поля 8, включаючи фронтальні очні поля.
Зв'язки й взаємодії
Префронтальна кора тісно взаємно пов'язана з більшістю структур мозку, включаючи особливо сильні зв'язки з іншими кортикальними, субкортикальними й стовбуровими утвореннями[9]. Дорсальна префронтальна кора найбільше поєднана з частинами мозку, що забезпечують увагу, когнітивну діяльність і моторику[10], в той час, як вентральна префронтальна кора взаємопов'язана з частинами мозку, що відповідають за емоції[11]. Префронтальна кора також має взаємні зв'язки зі стовбурною системою активації, і функціонування префронтальних регіонів сильно залежить від балансу активації/гальмування, що, відповідно до концепції трьох функціональних блоків А. Р. Лурії, показує взаємодії між першим, енергетичним, і третім, блоком програмування, регуляції й контролю психічної діяльності[12].
Медіальна префронтальна кора бере участь у генерації третьої й четвертої фази повільно-хвильового сну (ці фази об'єднуються під назвою «глибокий сон»), і її атрофія пов'язується зі скороченням частки глибокого сну щодо загального часу сну[13], що, відповідно, призводить до погіршення зберігання пам'яті.
Атрофія префронтальної кори відбувається природним чином у міру старіння, і було показано, що люди похилого віку мають проблеми з консолідацією пам'яті відповідно до деградації їхньої медіальної префронтальної кори. У літніх людей спогади замість передавання й збереження в неокортексі починають залишатися в гіпокампі, де вони були закодовані, це підтверджує підвищена гіпокампальна активація, на відміну від молодих дорослих індивідів, під час завдань на згадування інформації, коли випробовувані заучували словесні асоціації, спали й потім повинні були відтворити вивчені слова.
Функції
Базовою функцією префронтальної кори є комплексне керування розумовою й моторною активністю відповідно до внутрішніх цілей і планів[14].
Вона відіграє головну роль у створенні складних когнітивних схем і планів дій, прийнятті рішень, контролі й регуляції як внутрішньої діяльності, так і соціальної поведінки та взаємодії[15].
Функції керування префронтальної кори проявляються в диференціації суперечливих думок і мотивів та виборі між ними, диференціації та інтеграції об'єктів і понять, прогнозуванні наслідків справжньої активності та її коригування відповідно до бажаного результату, емоційної регуляції, вольового контролю, концентрації уваги на необхідних об'єктах[16].
Префронтальна кора — її дорсолатеральна частина — є також субстратом короткочасної пам'яті: Якобсен в 1936 році показав, що пошкодження префронтальної кори в приматів призводить до порушення функцій короткочасної пам'яті[17]; в 1952 році Карл Прібрам ідентифікував частину префронтальної кори, відповідальну за цей дефіцит, як 46 поле Бродмана, що також відоме як дорсолатеральна префронтальна кора[18]; пізніше, в 1993 році, Голдмен-Ракіч з колегами провела експеримент, де за допомогою тимчасової інактивації ділянок дорсолатеральної префронтальної кори вони викликали втрату спогадів, що зберігаються в короткочасній пам'яті[19].
Примітки
- Foundational Model of Anatomy
- Elisabeth Murray, Steven Wise, Kim Graham. Chapter 1: The History of Memory Systems // The Evolution of Memory Systems: Ancestors, Anatomy, and Adaptations. — 1st. — Oxford University Press, 2016. — С. 22—24. — ISBN 9780191509957.
- DeYoung C.G., Hirsh J.B., Shane M.S., Papademetris X., Rajeevan N., Gray J.R. Testing predictions from personality neuroscience. Brain structure and the big five // Psychological Science : journal. — 2010. — Vol. 21, no. 6 (6). — P. 820—828. — DOI:. — PMID:.
- Yang Y., Raine A. Prefrontal structural and functional brain imaging findings in antisocial, violent, and psychopathic individuals: a meta-analysis // Psychiatry Research : journal. — 2009. — Vol. 174, no. 2 (11). — P. 81—8. — DOI:. — PMID:.
- Miller E.K., Freedman D.J., Wallis J.D. The prefrontal cortex: categories, concepts and cognition // Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences : journal. — 2002. — Vol. 357, no. 1424 (8). — P. 1123—1136. — DOI:. — PMID:.
- Badre D., Kayser A.S., D'Esposito M. Frontal cortex and the discovery of abstract action rules // Neuron : journal. — Cell Press, 2010. — Vol. 66, no. 2 (4). — P. 315—326. — DOI:. — PMID:.
- Uylings HB, Groenewegen HJ, Kolb B. Do rats have a prefrontal cortex? // Behavioural Brain Research 146 (1-2). — 2003. — 9 листопада. — С. 3—17.
- Markowitsch HJ; Pritzel, M. The prefrontal cortex: Projection area of the thalamic mediodorsal nucleus? // Physiological Psychology 7 (1). — 1979. — 9 листопада. — С. 1—6.
- Alvarez JA, Emory E. Executive function and the frontal lobes: a meta-analytic review // Neuropsychology Review 16 (1). — 2006. — 9 листопада. — С. 17—42.
- Goldman-Rakic PS. Topography of cognition: parallel distributed networks in primate association cortex // Annual Review of Neuroscience 11. — 1988. — 9 листопада. — С. 137—156.
- Price JL. Prefrontal cortical networks related to visceral function and mood // Annals of the New York Academy of Sciences 877. — 1999. — 9 листопада. — С. 383—396.
- Лурия А. Р. Основы нейропсихологии. — 6-е изд. — Academia, 2008. — 384 с. — ISBN 978-5-7695-4915-1.
- Mander BA, Rao V, Lu B et al. Prefrontal atrophy, disrupted NREM slow waves and impaired hippocampal-dependent memory in aging // Nature Neuroscience 16 (3). — 2013. — 9 листопада. — С. 357—364.
- Miller EK, Freedman DJ, Wallis JD. The prefrontal cortex: categories, concepts and cognition // Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 357 (1424). — 2002. — 9 листопада. — С. 1123—1136.
- Yang Y, Raine A. Prefrontal structural and functional brain imaging findings in antisocial, violent, and psychopathic individuals: a meta-analysis // Psychiatry Research 174 (2). — 2009. — 9 листопада. — С. 81—88.
- Goldman-Rakic PS. The prefrontal landscape: implications of functional architecture for understanding human mentation and the central executive // Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 351 (1346). — 1996. — 9 листопада. — С. 1445—1453.
- Jacobsen C. F. Studies of cerebral function in primates. I. The functions of the frontal associations areas in monkeys // Comp Psychol Monogr 13. — 1936. — 9 листопада. — С. 3—60.
- Pribram, K. H.; Mishkin, M.; Rosvold, H. E.; Kaplan, S. J. Effects on delayed-response performance of lesions of dorsolateral and ventromedial frontal cortex of baboons // Journal of comparative and physiological psychology 45 (6). — 1952. — 9 листопада. — С. 565—575.
- Funahashi, S.; Bruce, C. J.; Goldman-Rakic, P. S. Dorsolateral prefrontal lesions and oculomotor delayed-response performance: Evidence for mnemonic "scotomas" // The Journal of neuroscience: the official journal of the Society for Neuroscience 13 (4). — 1993. — 9 листопада. — С. 1479—1497.
