Поле Бродмана 10
Поле Бродмана 10 (BA10) — одне з визначених Корбініаном Бродманом цитологічно визначених полів кори головного мозку. Це передня частина префронтальної кори в мозку людини.[2] BA10 було спочатку визначене Корбініаном Бродманом лише приблизно, виходячи з відмінностей цитоархітектоніки, які були помічені в трупних препаратах. Але з огляду на те, що навіть сучасні методики функціональної візуалізації також не можуть точно визначити межі поля, питання про його кордони залишається відкритим.
| Поле Бродмана 10 | |
|---|---|
 | |
 | |
| Деталі | |
| Ідентифікатори | |
| Латина | Area frontopolaris |
| NeuroNames | 76 |
| NeuroLex ID | birnlex_1741 |
| FMA | 68607[1] |
| Анатомічна термінологія | |
BA10 — найбільше з існуючих полів кори в людському мозку. Воно було описане як «одне з найменш зрозумілих регіонів людського мозку». дослідження припускає, що поле бере участь у стратегічних процесах пам'яті й різних виконавчих функціях. В ході еволюції, функції цієї ділянки в результаті розширилися по відношенню до іншої частини мозку.
Анатомія
Розмір
Обсяг BA10-близько 14 см3 і становить приблизно 1,2 % від загального обсягу об'єму мозку. Це вдвічі більше, ніж у людиноподібних з однаковим з людиною обсягом мозку. Для порівняння, обсяг BA10 у карликового шимпанзе становить близько 2,8 см3, і це лише 0,74 % від об'єму його мозку.
В кожній півкулі, площа 10 містить приблизно 250 мільйонів нейронів.
Розташування
BA10 є частиною визначеної цитоархітектонічно лобової ділянки кори головного мозку. Поле займає найбільш ростральну частину верхньої лобової звивини і середньої лобової звивини. В людському мозку, у медіальній частині півкулі вона обмежена знизу верхньою ростральною борозною. Вона не поширюється до поясної борозни. Цитоархітектонічно межує дорсально із полем Бродмана 9, каудально з полем Бродмана 46, а знизу з орбітальним полем Бродмана 47 і полем Бродмана 12.
Цитоархитектоніка
У шість шарів кори головного мозку в районі 10 були описані як такі, що виглядають «на диво, гомогенними». Всі вони легко ідентифікуються. По відношенню один до одного, шар І тонкий середній завширшки, займає становить 11 % від товщини поля 10. Другий шар тонкий і містить дрібні зернисті і пірамідальної середньо-темного забарвлення клітини при фарбуванні по Нісслю, яке зафарбовує РНК і ДНК. Найширший шар третій. Його пірамідальних нейронів менше, розташовані вони ближче один до одного, ніж у вищого шару II й нижчого шару ІV. Шари II і III складають 43 % товщі кори. Шар IV тонкий і має чіткі межі з сусідніми шарами III і V . Його клітини бліді, середнього зафарбування. Шар V широкий та складається з двох окремих підшарів, Vа і Vb. Щільність клітин Vа більше, ніж в Vb, підшар Vа має й темніше забарвлення. Шари IV і V складають 40 % товщини кори. Шар VI містить темні пірамідні і веретеноподібні нейрони. Він складає 6 % товщини Поля 10.
Поле 10 відрізняється від сусіднього Поля Бродмана 9 тим, що останній має більш виражений шар Vb і більш помітний шар II. Сусіднє Поле 11 Бродмана порівняно з Полем 10 має більш тонкий шар IV з більш помітними шарами Vа, Vb і II.
Поле 10 у людини має найнижчу щільність нейронів серед приматів. Також, його нейрони мають особливо великі дендритні розгалуження дуже щільно розташовані дендритні шипики. Вважається, що це уможливлює інтеграцію вхідної інформації з різних ділянок.
Структура поля
BA 10 розділене на три підполя, 10p, 10m і 10r. Підполе 10р займає фронтальний полюс, в той час як два інших накривають вентромедіальну частину префронтальної кори. Підполе 10м має тонкі шари II і IV і більш помітний шаром V. На противагу цьому, підполе 10р має помітний шар II і більш товстий шар IV. Великі пірамідальні клітини також присутні в підполі 10р. Шар III також великий. Але відзначається, що «відмінності між трьома підполями змазані й важко провести різкі межі між ними».
Зв'язки
Дослідження на приматах показали, що поле 10 контактує з іншми вищими асоціативнми ділянками кори, отримуючи вхідну й посилаючи вихідну інформацію. Зокрема, йдеться про префронтальну кору, первинну сенсорну й моторну ділянки. Через крайню капсулу поле пов'язане волокнами зі слуховою та мультисенсорною ділянками верхньої скроневої борозни. Волокна також продовжуються у медіальний поздовжній пучок, у білу речовинуі верхньої скроневої звивини (зони TAa, TS2, і TS3) і найближчі мультисенсорні зони на верхньому краї верхньої скроневої борозни (TPO). Ще одна область, пов'язана з цим полем через крайню капсулу - це вентральна ділянка острівцевої кори. Волокна до поясного пучка поєднують Поле 10 із передньою поясною корою, задньою поясною корою, і ретроспленальною корою (Поле Бродмана 26, Поле Бродмана 29 і Поле Бродмана 30). Гачкуватий пучок з'єднує його з мигдалеподібним тілом, темпорополярною корою і передньою частиною верхньої скроневої звивини. Поле не має зв'язків з тім'яною корою, потиличою корою, інтеротемпоральною корою.
В певних дослідженнях відзначено, що Поле 10 може бути анатомічною структурою для формування абстрактної обробки інформації та інтеграції результатів когнітивних операцій.
Еволюція
Катерина Семендефері (Katerina Semendeferi) зі співавторами вважає, що «в ході еволюції Поле 10 пройшло пару .. змін: одна включає в себе суттєве збільшення загального розміру поля, а інша - певне збільшення зв'язків особливо з іншими вищими асоціативними полями».
Дослідження рельєфу всередині черепа у Homo floresiensis виявило зростання в ділянці лобового полюсу, що може говорити про збільшення поля 10 Бродмана.
Функція
Хоча ця ділянка в людському мозку й велика, її функція є маловивченою. Koechlin & Hyafil запропонували вважати основною функцією фронтополярної кори когнітивний розподіл.[3] , що дозволяє раніше запущені завдання переводити у стані очікування для подальшої актуалізації й виконання після закінчення поточного завдання.
Багато які з наших складних форм поведінки і психічної діяльності вимагають одночасної участі декількох завдань. Дослідники припускають, що передня префронтальна кора може виконувати предметно-загальні функції плануванні складних паралельно-послідовних операцій. .[4] У 2006 році Мета-аналіз показав, що ростральна префронтальна кора задіяна в процесах робочої пам'яті, епізодичної пам'яті й координації паралельно-послідовного виконання завдань.[5]
Зображення
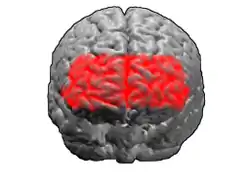
 Анімація.
Анімація.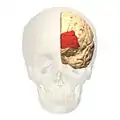 Фронтальний вигляд..
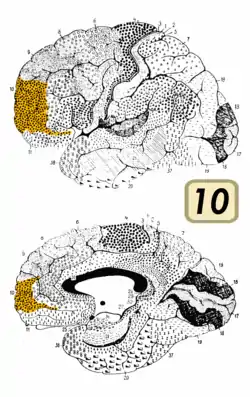
Фронтальний вигляд..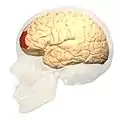 Латеральний вигляд.
Латеральний вигляд.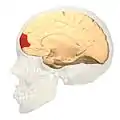 Медіальний вигляд.
Медіальний вигляд.
Див. також
Примітки
- Foundational Model of Anatomy
- Knowlton, Barbara J.; Morrison, Robert G.; Hummel, John E.; Holyoak, Keith J. (July 2012). A neurocomputational system for relational reasoning. Trends in Cognitive Sciences 16 (7): 373–381. PMID 22717468. doi:10.1016/j.tics.2012.06.002.
- Koechlin, E.; Hyafil, A. (2007). Anterior prefrontal function and the limits of human-decision making. Science 318: 594–598. doi:10.1126/science.1142995.
- Burgess, P.W.; Dumontheil, I.; Gilbert, S.J. (2007). The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Science 11: 7.
- Gilbert, Sam J.; Spengler, Stephanie; Simons, Jon S.; Steele, J. Douglas; Lawrie, Stephen M.; Frith, Christopher D.; Burgess, Paul W. (1 червня 2006). Functional specialization within rostral prefrontal cortex (area 10): a meta-analysis. Journal of Cognitive Neuroscience 18 (6): 932–948. ISSN 0898-929X. PMID 16839301. doi:10.1162/jocn.2006.18.6.932.
Посилання
- Поле Бродмана 10
- BrainMaps, Поле Бродмана 10 Префронтальної Кори
- Поле Бродмана 10, ключ до еволюції людини?