Виродження (біологія)
У біологічних системах виродження відбувається, коли структурно відмінні компоненти/модулі/шляхи можуть за певних умов виконувати подібні функції (тобто фактично взаємозамінні), але виконувати різні функції в інших умовах.[1] [2] Таким чином, виродженість є властивістю відношення, яка вимагає порівняння поведінки двох або більше компонентів. Зокрема, якщо виродженість присутня в парі компонентів, то існуватимуть умови, коли пара буде виглядати функціонально зайвою.[1] [3]
Зауважте, що таке використання терміну практично не має відношення до сумнівно змістовної концепції дегенерації популяцій (деволюції), які втратили функції предків.
Біологічні приклади
Приклади виродження можна знайти в генетичному коді, коли багато різних нуклеотидних послідовностей кодують один і той же поліпептид; у згортанні білків, коли різні поліпептиди є структурно та функціонально еквівалентними; у функціях білка, коли спостерігаються перехресні функції зв'язування, і подібні каталітичні специфічності; в метаболізмі, коли можуть співіснувати множинні паралельні шляхи біосинтезу та катаболізму. Загалом, виродження спостерігається в білках кожного функціонального класу (наприклад, ферментативних, структурних або регуляторних),[4] [5] білкових комплексах, [6] онтогенезі, [7] нервовій системі, [8] передачі сигналів у клітинах (перехресні перешкоди) та в численних інших біологічних системах.[1]
Внесок у міцність
Виродження сприяє стійкості біологічних ознак за допомогою кількох механізмів. Вироджені компоненти компенсують один одного в умовах, коли вони є функціонально надлишковими, забезпечуючи тим самим стійкість проти збоїв компонента або шляху. Оскільки вироджені компоненти дещо відрізняються, вони, як правило, володіють унікальною чутливістю, тому цілеспрямована атака, така як специфічний інгібітор, менш ймовірно несе ризик для всіх компонентів одночасно.[3] Є численні біологічні приклади, коли виродження таким чином сприяє міцності. Наприклад, сімейства генів можуть кодувати різноманітні білки з багатьма відмінними ролями, але іноді ці білки можуть компенсувати один одного під час втраченої або пригніченої експресії генів, як видно з ролей розвитку сімейства генів адгезинів у Saccharomyces.[9] Поживні речовини можуть метаболізуватися різними метаболічними шляхами, які ефективно взаємозамінні для певних метаболітів, навіть якщо загальний результат кожного шляху не ідентичний.[10] [11] При злоякісних пухлинах, методи лікування, орієнтовані на рецептор EGF обходяться ко-активацією альтернативних рецепторних тирозинкіназ (РСІ), які мають часткове функціональне перекриття з рецептором EGF (і, отже, виродженими), але не є мішенню специфічного інгібітора рецептора EGF.[12] [13] Інші приклади з різних рівнів біологічної організації можна знайти в [1]
Теорія
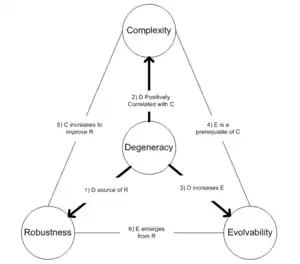
Кілька теоретичних розробок окреслили зв’язки між виродженням і важливими біологічними мірами, пов’язаними зі стійкістю, складністю та можливістю еволюціонувати. До них належать:
- Теоретичні аргументи, підтверджені моделюванням, припускають, що виродження може призвести до розподілених форм надійності в мережах взаємодії білків.[14] Ці автори припускають, що подібні явища, ймовірно, виникнуть в інших біологічних мережах і потенційно можуть також сприяти стійкості екосистем.
- Тононі та ін. знайшли докази того, що виродження невіддільне від існування ієрархічної складності в нейронних популяціях.[8] Вони стверджують, що зв'язок між виродженням і складністю, ймовірно, буде набагато більш загальним.
- Досить абстрактне моделювання підтвердило гіпотезу про те, що виродження фундаментально змінює схильність генетичної системи отримувати доступ до нових спадкових фенотипів [15] і що виродження може бути передумовою еволюції з відкритим фіналом.
- Три вищенаведені гіпотези були інтегровані в [3] де припускається, що виродження відіграє центральну роль у еволюції біологічної складності з відкритим фіналом. У тій же статті стверджувалося, що відсутність виродження в багатьох розроблених (абіотичних) складних системах може допомогти пояснити, чому надійність, здається, суперечить гнучкості та адаптивності, як це видно в програмному забезпеченні, системній інженерії та штучному житті . [3]
Дивіться також
- Каналізація
- Еквіфінальність системи
Посилання
- Edelman and Gally; Gally, J. A. (2001). Degeneracy and complexity in biological systems. Proceedings of the National Academy of Sciences, USA 98 (24): 13763–13768. Bibcode:2001PNAS...9813763E. PMC 61115. PMID 11698650. doi:10.1073/pnas.231499798. Проігноровано невідомий параметр
|doi-access=(довідка) - Mason, Paul H. (2 січня 2015). Degeneracy: Demystifying and destigmatizing a core concept in systems biology. Complexity 20 (3): 12–21. Bibcode:2015Cmplx..20c..12M. doi:10.1002/cplx.21534.
- Whitacre (2010). Degeneracy: a link between evolvability, robustness and complexity in biological systems. Theoretical Biology and Medical Modelling 7 (6): 6. Bibcode:2009arXiv0910.2586W. PMC 2830971. PMID 20167097. arXiv:0910.2586. doi:10.1186/1742-4682-7-6.
- Atamas (2005). Les affinités électives. Pour la Science 46: 39–43.
- Wagner (2000). The role of population size, pleiotropy and fitness effects of mutations in the evolution of overlapping gene functions. Genetics 154 (3): 1389–1401. PMC 1461000. PMID 10757778. doi:10.1093/genetics/154.3.1389.
- Kurakin (2009). Scale-free flow of life: on the biology, economics, and physics of the cell. Theoretical Biology and Medical Modelling 6 (1): 6. PMC 2683819. PMID 19416527. doi:10.1186/1742-4682-6-6.
- Newman (1994). Generic physical mechanisms of tissue morphogenesis: A common basis for development and evolution. Journal of Evolutionary Biology 7 (4): 480. doi:10.1046/j.1420-9101.1994.7040467.x. Проігноровано невідомий параметр
|doi-access=(довідка) - Tononi; Sporns, O.; Edelman, G. M. (1999). Measures of degeneracy and redundancy in biological networks. Proceedings of the National Academy of Sciences, USA 96 (6): 3257–3262. Bibcode:1999PNAS...96.3257T. PMC 15929. PMID 10077671. doi:10.1073/pnas.96.6.3257. Проігноровано невідомий параметр
|doi-access=(довідка) - Guo; Styles, C. A.; Feng, Q.; Fink, G. R. (2000). A Saccharomyces gene family involved in invasive growth, cell-cell adhesion, and mating. Proceedings of the National Academy of Sciences, USA 97 (22): 12158–12163. Bibcode:2000PNAS...9712158G. PMC 17311. PMID 11027318. doi:10.1073/pnas.220420397. Проігноровано невідомий параметр
|doi-access=(довідка) - Kitano (2004). Biological robustness. Nature Reviews Genetics 5 (11): 826–837. PMID 15520792. doi:10.1038/nrg1471.
- Ma and Zeng; Zeng, AP (2003). The connectivity structure, giant strong component and centrality of metabolic networks. Bioinformatics 19 (11): 1423–1430. PMID 12874056. doi:10.1093/bioinformatics/btg177. Проігноровано невідомий параметр
|doi-access=(довідка) - Huang; Mukasa, A.; Bonavia, R.; Flynn, R. A.; Brewer, Z. E.; Cavenee, W. K.; Furnari, F. B.; White, F. M. (2007). Quantitative analysis of EGFRvIII cellular signaling networks reveals a combinatorial therapeutic strategy for glioblastoma. Proceedings of the National Academy of Sciences 104 (31): 12867–72. Bibcode:2007PNAS..10412867H. PMC 1937558. PMID 17646646. doi:10.1073/pnas.0705158104. Проігноровано невідомий параметр
|doi-access=(довідка) - Stommel; Kimmelman, AC; Ying, H; Nabioullin, R; Ponugoti, AH; Wiedemeyer, R; Stegh, AH; Bradner, JE та ін. (2007). Coactivation of receptor tyrosine kinases affects the response of tumor cells to targeted therapies. Science 318 (5848): 287–90. Bibcode:2007Sci...318..287S. PMID 17872411. doi:10.1126/science.1142946.
- Whitacre and Bender; Bender, Axel (2010). Networked buffering: a basic mechanism for distributed robustness in complex adaptive systems. Theoretical Biology and Medical Modelling 7 (20): 20. PMC 2901314. PMID 20550663. doi:10.1186/1742-4682-7-20.
- Whitacre and Bender; Bender, A (2010). Degeneracy: a design principle for achieving robustness and evolvability. Journal of Theoretical Biology 263 (1): 143–153. Bibcode:2010JThBi.263..143W. PMID 19925810. arXiv:0907.0510. doi:10.1016/j.jtbi.2009.11.008.