Нервова система
Нервова система — цілісна морфологічна і функціональна сукупність різних взаємопов'язаних нервових структур тварин та людей, яка спільно з гуморальною системою забезпечує взаємопов'язану регуляцію діяльності усіх систем організму та реакцію на зміну умов внутрішнього та зовнішнього середовища. Нервова система діє як інтегративна, зв'язуючи в єдине ціле чутливість, рухову активність та роботу інших регуляторних систем, наприклад, як ендокринна та імунна системи.
| Нервова система | |
|---|---|
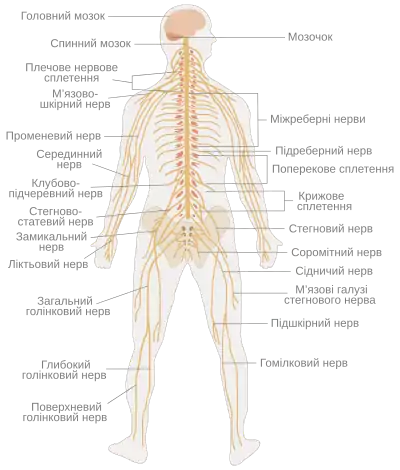 Нервова система людини | |
| Деталі | |
| Ідентифікатори | |
| Латина | systema nervosum |
| MeSH | D009420 |
| TA98 | A14.0.00.000 |
| FMA | 7157[1] |
| Анатомічна термінологія | |
У більшості тварин нервова система складається з двох частин — центральної та периферичної. Центральна нервова система хребетних (зокрема людини) складається з головного та спинного мозку. Периферична нервова система складається з сенсорних нейронів, сукупностей нейронів, що називаються ганглії, та нервів, що сполучають їх між собою та з центральною нервовою системою.
Нерви залежно від складу їхніх волокон поділяють на чутливі, рухові і змішані. Чутливі нерви містять доцентрові волокна, рухові — відцентрові волокна, а змішані — обидва види нервових волокон. Багато нервів та їхніх розгалужень на периферії крім нервових волокон мають нервові вузли (ганглії). Вони складаються з нейронів, відростки яких входять до складу нервів, та їхніх розгалужень (нервові сплетення).
Загальні риси розвитку нервової системи
Розвиток у безхребетних
У безхребетних нервова система розвивається з ектодерми, яка під дією специфічних факторів перетворюється у спеціальну ектодерму — нейроектодерму. Ектодерма, з якої походитиме нервова система розміщена вентрально, тобто спереду.[2]
Сам процес утворення нейроанатомічних утворів в багато чому подібний, хоча має певні відмінності серед різних класів. Найбільше процеси розвитку нервової системи вивчений на C. elegans та дрозофілах. Так у C. elegans попередники нервових клітин занурюються вглиб ембріона, де опісля утворюють нейрони та гліальні клітини.[2] У дрозофіл попередники нервових клітин спочатку розташовані збоку, опісля мігрують до середньої лінії та занурюються всередину ембріона (деламінація). Оскільки дрозофіла є сегментованим організмом, то у кожному сегменті відбуваються подібні процеси деламінації, які ведуть до утворення черевного ланцюжка, що об'єднує нервові вузли, та периферичних відгалужень.[2][3]
Розвиток у хребетних
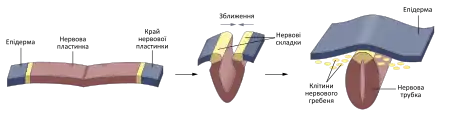
У хребетних нервова система також розвивається з ектодерми, однак, ектодерма, з якої розвинеться нервова система розміщена дорсально (ззаду) і над нотохордою[4][5].
Спочатку ця ектодерма потовщується і утворюється нервова пластинка — утвір, який дасть початок і ЦНС, і ПНС. Далі відбувається явище, яке отримало назву нейруляції — перетворення нервової пластинки у нервову трубку. Нейруляція у хребетних буває первинною та вторинною.[3][4] Первинна нейруляція являє собою сходження країв нервової трубки — нервових складок. Нервові складки виникають внаслідок того, що краї нервової пластинки розростаються, а в самій нервовій пластинці утворюється заглиблення — нервова борозна. Нервові складки сходяться та надають порожнистої структури нервовій трубці.[3][6] Нервова трубка втрачає контакт з ектодермою, яка дасть початок шкірним покривам, і повністю занурюється в товщу ембріона[7]. При цьому найбільш бічна ділянка нейроектодерми, яка контактує з ектодермою, яка утворюватиме шкірні покриви, дає початок праобразу ПНС — нервовому гребеню[8], а у краніальній ділянці ще й нервовим плакодам[9][10]. З цих утворів виникатимуть нейрони чутливих та вегетативних вузлів, периферична глія, а також ряд ненервових утворів (мозкова речовина надниркових залоз, наприклад)[11]. При вторинній нейруляції мезенхімальні клітини утворюють частину спинного мозку. Згодом в цій частині виникають декілька порожнин, які зливають та утворюють таку ж порожнину, як утворюється при первинній нейруляції. Порожнина від первинної нейруляції зливається з порожниною від вторинної і утворюється єдина порожнина.[12][13]
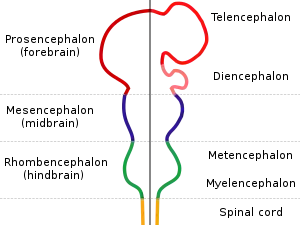
У різних класів різні ділянки нервової трубки утворюються первинною або вторинною нейруляцією; в основному передня і більша частина утворюється первинною нейруляцією, а задня — вторинною[6].
Незабаром після утворення нервової трубки її потовщена передня ділянка дає початок трьом мозковим міхурцям, які притаманні всім хребетним: ромбоподібному (первинному задньому) мозку, середньому мозку та передньому мозку. В свою чергу три первинних мозкових міхурці дають початок п'ятьом вторинним: передній мозок ділиться на кінцевий (дасть початок півкулям кінцевого мозку і їх утворам) та проміжний (дасть початок однойменному мозку), а ромбоподібний — на задній (дасть початок мосту та мозочку) та довгастий (дасть початок однойменному мозку). З решти нервової трубки виникає спинний мозок.[14][15]
Нервова тканина
Нервова тканина є основою нервової системи та складається з двох видів клітин — нейронів та нейроглії. Перші клітини виконують специфічні для нервової системи завдання (проведення нервових імпульсів, аналіз інформації, трофічна функція), а другі — всіляко забезпечують оптимальні умови для існування та функціонування нейронів.
Нейрон та його відростки
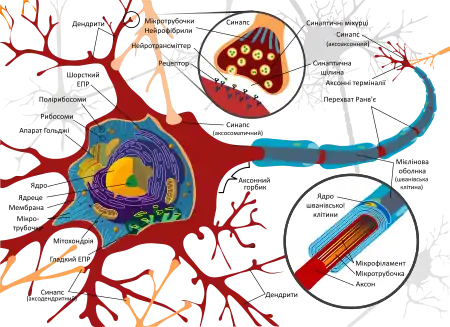
Нейрон, або нейроцит, є збудливою клітиною, яка сприймає та обробляє інформацію, що надходить з внутрішнього або зовнішнього середовища та, генеруючи потенціал дії, надсилає імпульси для відповіді на зміни у цих середовищах. Нейрони мають практично усі багатоклітинні організми (серед винятків, наприклад, губки та пластинчасті).
Нейрон складається з тіла (навколоядро, або перикаріон)[16] та відростків — дендритів, яких зазвичай є багато і які сильно галузяться, утворюючи дендритне дерево, та аксона, найдовшого відростка, який завжди є один, але може давати колатералі різної довжини.[16][17][18] Дендрити деяких клітин (наприклад, клітин Беца) мають своєрідні вип'ячування — дендритні шипики, які дозволяють збільшувати контактну площу клітини; у дендритів інтернейронів шипики практично відсутні[17][19]. Кількість шипиків не є постійною і міняється протягом життя. По дендритах сигнал завжди прямує до тіла, або аферентно[16][18]. Початковий сегмент аксона — аксонний горбик — є ключовим в генерації потенціалу дії[20]. По аксону сигнал прямує від тіла нейрона, тобто еферентно. В кінці аксон розгалужується на терміналі, які утворюють пресинаптичну мембрану синапса.[16][18] Окрім того, існують нейрони з третім типом відростків – амакриновими відростками. По цих відростках імпульс може прямувати в обидві сторони.[16][21] Існують також нейрони, які не мають дендритів, але мають аксон.[22]
Є ряд характерних знахідок в цитоплазмі, які характеризують функції нейрона. Так, у цитоплазмі знаходять велику кількість мітохондрій, що вказує на високу метаболічну активність нейрона[18]. Характерною ознакою нейрона є речовина Ніссля, або базофільна речовина, яка є скупченням гранулярного ендоплазматичного ретикулуму. Це свідчить про високий рівень процесів синтезу білків в нейронах.[19] В ядрі переважає еухроматин, що теж свідчить про активність транскрипції. Як для цитоплазми перикаріона, так і для відростків характерною ознакою є добре розвинутий цитоскелет, який представлений мікротрубочками та нейрофіламентами, і який пов'язаний з процесами транспорту та стабілізації просторової будови відростків та тіла, компартментилізації цитоплазми[17].
Нейроглія
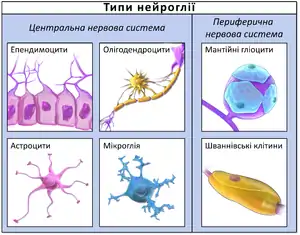
Під нейроглією, або просто глією, розуміють сукупність клітин, різних за походженням, функції яких зводяться до одного — підтримання нормального функціонування нейронів, а отже – нервової системи[23]. У людини клітини нейроглії за своєю кількістю значно переважають кількість нейронів[24]. Розрізняють макроглію, яка має ектодермальне походження (таке саме походження, як і нейрони) та мікроглію, яка має мезодермальне походження[25][26]. Також нейроглію у ссавців ділять за анатомічним принципом на нейроглію ЦНС та нейроглію ПНС. До нейрогії ЦНС відносять астроглію, олігодендритоглію, NG-2-глію та мікроглію. До нейроглії ПНС відносять шваннівські клітини, мантійні гліоцити, ентеричну глію та нюхові оболонкові клітини.[25]
Їхні функції різняться: шваннівські клітини та олігодендроцити утворюють мієлін в ПНС та ЦНС відповідно, астроцити є ключовими в регулюванні гомеостазу, мікроглія забезпечує фагоцитоз, ентерична глія, мантійні гліоцити та нюхові оболонкові клітини відповідають за підтримку гомеостазу в ентеричній нервовій системі, нервових вузлах та нюховій цибулині відповідно. Вище перелічені функції є лише основними, кожний тип глії володіє значно більшим діапазоном функцій (наприклад, мікроглія окрім фагоцитозу є дуже важливою у процесі розвитку нервової системи, синаптичній передачі та старінні).
Нейроглія наявна не тільки у ссавців, а й у більшості інших тварин. Так, глія відсутня у кнідарій і реброплавів. У ацеломорфів уже наявні клітини, функції яких нагадують функції глії. Клітини з подібними функціями знайдені у деяких плоских червів. У круглих червів наявні протоастроцити.[27] У кільчаків нейроглія ускладнюється: наявні гігантські гліальні клітини, функції яких подібні до астроцитів, гліальні клітини, які оточують перикаріон, та гліальних клітин, які оточують аксони[28]. В подальшому від типу до типу і від класу до класу відбувається ускладнення та збільшення різноманітності гліальних клітин[27].
Анатомічно-фізіологічний поділ та основні принципи будови
Нервову систему ділять за анатомічно-фізіологічним принципом. Слід зауважити, що такий поділ не позбавлений умовностей. Так, анатомічний поділ на центральну та периферичну нервові системи не зберігає функціональну єдність нервової системи (наприклад, нервовий шлях має складові, які знаходяться як в ЦНС, так і в ПНС).[29] Цього позбавлений поділ нервової системи на соматичну та вегетативну. Такий поділ, як і анатомічний, є дещо умовним, проте також широко використовується.[30][31]
Анатомічний поділ та основні принципи в будові
У більшості тварин, за винятком тих, у яких наявна дифузна нервова система, анатомічно виділяють центральну та периферичну частини, або центральну (ЦНС) та периферичну (ПНС) нервові системи. Якщо взяти хребетних, то у них центральна нервова система, складається з головного мозку та спинного мозку.[29] Головний мозок у свою чергу ділиться на ряд менших частин, які мають специфічну будову та забезпечують специфічні функції: довгастий мозочок, міст (тільки у ссавців і, можливо, у птахів), мозочок, середній мозок, проміжний мозок та кінцевий мозок.[32] Від головного мозку відходять черепні нерви, а від спинного – спинномозкові нерви. Ці дві групи нервів разом з нервовими вузлами та нервовими сплетеннями утворюють периферичну нервову систему.[29][33]
У безхребетних центральна нервова система складається з нервових вузлів та нервових стовбурів, які об'єднують нервові вузли в одне ціле. Від вузлів відходять нерви, які безпосередньо іннервують органи-мішені (органи чуття та решту внутрішніх органів), які формують периферичну нервову систему безхребетних.[34]

Фізіологічний поділ нервової системи
Нервову систему у функціональному плані прийнято ділити на соматичну (анімальну) та вегетативну (автономну). Першу пов'язують з виконанням свідомих дій, тоді як другу — з несвідомими процесами, які перебігають в організмі живої істоти та які покликані підтримувати гомеостаз та цілісність організму.[29][31] І перша, і друга системи мають аферентні ланки, які пов'язані з рецепторами. Третя система, завдяки якій нервова система керує тілом — це нейроендокринна система.[35] Так, класично вважається, що парасимпатична та симпатична нервові системи перебувають в антагоністичних стосунках. Проте, згідно з сучасними уявленнями, вони перебувають не в суто антагоністичних стосунках, а радше в різних комбінаціях активуються між собою в залежності від потреб організму, хоча для зручності їхні дії визначають як антагоністичні.[36] Вважається, що у ссавців симпатична нервова система пришвидшує серцебиття, розширює бронхи та зіниці, сповільнює перистальтику ШКТ, а парасимпатична має протилежну дію.[37]
У ссавців вегетативну систему прийнято ділити на симпатичну, парасимпатичну та ентеричну. Симпатична та парасимпатична нервові системи мають центральну та периферичну складові.[38][39] Центральна симпатична нервова система розміщена в грудній та поперековій частинах, в бічних рогах спинного мозку. Центральна парасимпатична нервова система розміщена в деяких ядрах III, VII, IX та X пар черепних нервів та у крижовому відділі спинного мозку. Периферичні частини обох цих систем розміщені у вегетативних вузлах; нейрони цих вузлів безпосередньо іннервують орган-мішень. Головним вегетативним і нейроендокринним центром в ЦНС вважають гіпоталамус.[39][40][30]
Ентерична нервова вважається окремим відділом вегетативної нервової системи, оскільки вона має власний рефлекторний апарат (містить чутливі нейрони та інтернейрони)[39][41].
У інших хребетних вегетативна нервова система побудована за подібним принципом. Центральна вегетативна нервова система ділиться на черепну частину, яка є парасимпатичною, та хребетну, в якій поділ на симпатичну чи парасимпатичну складову провести не можна. Симпатичні стовбури присутні практично у всіх хребетних, окрім круглоротих, у яких ці вузли або відсутні, або розкидані по тілу, та пластинозябрових (підклас хрящових риб), у яких від неповний. Ентерична нервова система наявна у всіх хребетних і знаходиться під контролем ЦНС.[42]
У безхребетних та частина нервової системи, яка іннервує органи, називається вісцеральною нервовою системою. В ній немає периферичної частини, а контроль за внутрішніми органами перебирають на себе нервові вузли. Як і у хребетних поведінка безхребетних визначається тісною взаємодією вісцеральної та соматомоторної нервових систем. Окрім того, як і у хребетних, у безхребетних наявні ритмічні повторювані функції, такі як серцебиття чи перестальтика, які теж забезпечуються вісцеральною нервовою системою.[43]
Тенденції еволюції нервової системи та її порівняльна анатомія
Кнідарії та реброплави
Нервова система, яка наявна у кнідарій та реброплавів (утворюють невизначений таксономічний ранг кишковопорожнинних) називається дифузною нервовою системою[44]. Згідно з класичними уявленнями така нервова система характеризується абсолютною децентралізацією — нейрони утворюють нервову сітку по всьому тілі тварин[45][46]. Однак, навіть в ній можна простежити тенденції до централізації та цефалізації[21][47].
У реброплавів нервова система складається зі субепідермального сплетення, яке компактно розташоване вздовж гребних пластинок ребер та біля рота. Проте найбільше нервових клітин міститься біля аборального органу. Аборальний орган є центром балансу та орієнтації. Він складається зі статоліта (вапнякова кулька), яка розміщена на чотирьох дужках. Під зміною тиску дужки прогинаються. Від кожної дужки відходить пара борозенок, яка іннервує гребні пластинки певного квадранта та регулює рух тварини.[48]
У кнідарій зазвичай наявні два нервових сплетення: субепідермальне та субгастродермальне; останнє розвинуте гірше. Загалом розвиток нервової системи та органів чуття залежить від способу життя: у поліпів нервова система розвинута гірше, ніж у вільноживучих осіб. У деяких медуз наявні додаткові сплетення.[46] У багатьох сцифоїдних медуз та гідр спостерігається тенденція до виникнення нервових кілець, які ще краще дозволяють тварині реагувати на подразнення та є ознаками централізації[49][50][51]. Серед утворів, які тяжіють до централізації також є «гігантські аксони», які за своєю природою є синцитіями[52][47]. Через наявність таких утворів деякі вчені виділяють в цих тварин ЦНС та ПНС[53]. Серед органів чуття розрізняють статоцисти та світлочутливі вічка; у кубомедуз наявні добре розвинуті очі[51]. Окрім нервової системи є і ненервові утвори, які реагують на подразники — так звані незалежні ефектори. До них належать неманоцити та збудливі епітеліальні клітини.[52][49]
Незважаючи на свою простоту нервова система кнідарій та реброплавів вже має ряд ознак, які характерні в будові та функціонуванні інших типів нервових систем та дозволяє цим тваринам активно шукати їжу, пересуватися в її напрямку, реагувати на подразники. Так, у цих тварин є чутливі та рухові нейрони (є також невелика кількість сенсорно-моторних нейронів).[54] Чутливі нейрони сприймають подразнення і передають збудження на руховий нейрон, який в свою чергу іннервує орган-мішень. Багато мотонейронів мають окрім аксона також амакринові відростки і, таким чином, передають імпульс один одному, що значно пришвидшує відповідь на подразник.[21] У функціональному плані для цієї нервової системи характерні фундаментальні закони передачі імпульсу (від чутливого нейрона до рухового, від дендрита до аксона), конвергенції та дивергенції[21], наявні хімічні синапси[47] (а у гідроїдних знайдені ще й електричні синапси[55]). Зважаючи на наявність амакринових відростків більшість хімічних синапсів є симетричними — тобто імпульс через ці сигнали передається в обидвох напрямках[55].
Плоскі черви
Нервова система плоских червів піддається трьом основним тенденціям її еволюційного розвитку: централізації, симетрії (плоскі черви — білатеральні тварини) та цефалізації. У плоских червів наявні скупчення нейронів, які називають вузлами. Таке скупчення дозволяє значно зменшувати час взаємодії між нейронами. Найбільшими вузлами є парні головні вузли, які локалізуються в ростральній (передній кінець) частині черва (простежується цефалізація та симетрія).[21][34][56] Ці вузли вважають справжнім головним мозком (але це твердження підтримують не всі вчені), який контролює рефлекторну діяльність інших відділів нервової системи. Кількість нейронів в цих вузлах варіює в залежності від виду та способу життя черва.[34][57] Так, у вільноживучих червів розвиток і нервової системи, і органів чуття кращий, ніж у паразитичних плоских червів[34].
Від головних вузлів відходять поздовжні нервові стовбури (утворені аксонами), які об'єднують разом головні вузли та інші менші нервові вузли (простежується централізація).[21][34] Кількість цих поздовжніх стовбурів різниться у різних видів; чим складніша поведінка черва, тим менше нервових стовбурів є у нього, що теж свідчить на користь централізації та конденсації нервових утворів[56]. Зазвичай є три парні стовбури: найбільший передній, задній та бічний[58]. Поздовжні стовбури об'єднані один з одним завдяки поперечним (кільцевим) нервовим стовбурам, кількість яких теж неоднакова у різних видів. Наявність поздовжніх та поперечних стовбурів нагадують драбину, саме тому плоскі черви є тваринами з драбинчастою нервовою системою.[34][57]
Вузли та нервові стовбури утворюють ЦНС. До складу ПНС входять відгалуження від вузлів, а також нервові сітки, які ще зберігаються у цих тварин.[56][34]
У плоских червів вперше з'являються інтернейрони. Вони можуть бути збуджуючими або гальмівними. Як і мотонейрони вони в розташовані в нервових вузлах. На відміну від кнідарій та реброплавів основна кількість мото- та інтеронейронів є уніполярними. Чутливі нейрони є біполярними. Синапси в основному утворюються в товщі вузла (від аксонів відходять декілька відгалужень, які виконують роль дентритів).[59]
Наявність глії в плоских червів та інших близьких типів є дискутабельним питанням[57]. Гліальні, або підтримуючі клітини відсутні в таких рядах плоских червів, як Catenulida та Macrostomida. З іншого боку такі клітини наявні в таких рядах плоских червів, як Policladida і в деяких Tricladida.[60]
Круглі черви: глія та нервова система C. elegans
У нематод будова нервової системи типова, хоча відмінності між видами наявні. Нервова система круглих червів нагадує драбинчасту нервову систему плоских червів. Так, характерним утвором нервової системи круглих червів є навкологлоткове кільце. Навкологлоткове кільце складається з комісур, які об'єднують парні передні та бічні вузли та непарний задній вузол в один єдиний комплекс. Через своє розташування довкола глотки це кільце і отримало свою назву. В напрямку до голови відходять нерви до органів чуття. Від навкологлотково кільця відходять нервові стовбури (в різних видів їх від чотирьох і більше). Найкраще розвинутим є передній нервовий стовбур, який по суті є ланцюжком нервових вузлів. Передній нервовий стовбур простягається аж до ділянки відхідника. Решту нервових стовбурів в основному складаються з тільки з нервових волокон. Як і у плоских червів, у вузлах розміщені інтер- та мотонейрони. Клітини, які виконують підтримуючу функцію щодо нейронів, в основному пов'язані з чутливими нейронами та такими органами чуття, як сенсили.[34][61][62]
Зважаючи на простоту нейроанатомії, нематоди стали улюбленими тваринами для досліджень пов'язаних з нервовою системою. Особливо дослідженим є черв Caenorhabditis elegans. У кожної особини гермафродита (генотип XX) наявно 302 нейрона та 56 гліальних клітин[60]. Така кількість відносна простота дозволила ставити на цьому черв'яку велику кількість дослідів, які стосують ембріонального розвитку нейронів, їх міграції, дослідження комунікацій між нейронами, тощо.
Кільчаки
Кільчаки мають одну особливість — їхнє тіло є сегментованим. Кожен сегмент є в чомусь подібний на інший. Сегментарний апарат впливає на будову нервової системи, яка тепер теж має сегментарну будову, тобто частини, які подібні і повторюються.[22] Таку тенденцію можна простежити і у значно складніших у будові тварин, наприклад, у хребетних (сегменти спинного мозку).
Головний мозок (головний нервовий вузол, або надглотковий вузол) може розміщуватися в найпершому сегменті (у випадку багатощетинкових червів[63][64]), або перебувати ближче до задньої частини (у випадку п'явок та малощетинкових червів)[65][66][67]). Завдяки навкологлотковому кільцю мозок з'єднується з підглотковим вузлом та черевним нервовим ланцюжком, який може утворюватися злиттям двох передніх нервових стовбурів у малощетинкових червів[64][62], або бути представленим двома нервовими стовбурами у випадку багатощетинкових червів[64]. У цих стовбурах наявні гігантські аксони.[63][65]. Сам ланцюжок об'єднує нервові вузли. Оскільки у багатощетинкових червів нервових ланцюжків двоє, то й на кожен сегмент припадає два нервові вузли, які з'єднані поперечними волокнами. Така нервова система нагадує драбинчасту нервову систему, подібну до нервової системи плоских червів.[64] Якщо нервовий стовбур один, то на кожен сегмент зазвичай припадає один об'єднаний нервовий вузол і така нервова система належить до вузлового типу.[67][62]. До того ж простежується тенденція до їх злиття, як, наприклад, у випадку з п'явками.[28] Від вузлів відходять відростки, які формують периферичну нервову систему, іннервують рецептори (у цьому випадку радше від рецепторів волокна прямують до вузла) та м'язи[62].
Членистоногі
Нервова система у членистоногих також має типову будову для всіх класів, але її рівень розвитку залежить від складності розвитку нервової системи і вона в багато чому подібна на нервову систему кільчаків[68][69].
Доброго розвитку зазнає головний мозок (надглотковий вузол), який утворений злиттям нервових вузлів. У більшості членистоногих у ньому розрізняють три частини[68][70][71], а у хеліцерових та морських павуків — дві, бо у них відсутній дейтоцеребрум[72][73]:
- протоцеребрум — передня частина головного мозку, яка складається з зорових часток (центр аналізу зорової інформації), присереднього центрального комплексу, який є руховим центром, та грибоподібних тіл, які відсутні в ракоподібних, і пов'язані з нюховою інформацією та навчанням;
- дейтероцеребрум — середня частина головного мозку, невромер першго антенального сегмента, який пов'язаний з аналізом нюхової інформації;
- тритоцеребрум — є найменшою частиною мозку членистоногих, невромером другого антенального сегмента; іннервує нижню частину голови, утворює навкологлоткове кільце та єднає завдяки цим комісурам головний мозок з підглотковим нервовим вузлом та черевним нервовим стовбуром (або нервовими стовбурами).
Останній також утворився внаслідок об'єднання декількох нервових вузлів (зазвичай чотирьох-п'яти нервових вузлів) та іннервує органи ротової порожнини, слинні залози, локально розташовані м'язи[68][74].
Нервові стовбури та пов'язані з ними вузли у різних видів можуть бути розділеними, або зливатися в одне. Їхнє злиття залежить від будови тіла комахи: якщо у комахи простежується об'єднання окремих сегментів (тагм), то і нервова система буде слідувати цьому[68][75]. Так, у багатьох зяброногих раків, ремипедій, цефалокарид, деяких примітивних безкрилих комах і черевні нервові стовбури, і нервові вузли залишаються розділеними; вузли об'єднані між собою завдяки поперечним комісурам[69][76]. У більшості багатоніжок нервові стовбури розділені, а в кожному сегменті наявні об'єднані нервові вузли; у двопарноногих у кожному сегментів наявні дві пари об'єднаних нервових вузлів[74]. У решти членистоногих і нервові стовбури, і нервові вузли зливаються у різних пропорціях між собою[69]. Це яскраво простежується у хеліцерових: у деяких павуків не можливо простежити сегментарність у будові нервової системи, оскільки нервові вузли головогрудей та черевця формують єдину нервову масу[72]. Те саме відбувається у десятиногих: наприклад, у креветок грудні та черевні нервові вузли зливаються між собою в поперечному напрямку, але розділені у поздовжньому напрямку; у крабів черевні та грудні вузли формують один єдиний нервовий конгломерат[77]. Прикладом таких процесів серед комах є двокрилі, у яких черевні нервові вузли з'єднаються в одне ціле[76].
Фізіологія
Потенціал спокою
В спокійному стані внутрішня поверхня плазмалеми є негативно зарядженою, саме тому потенціалом спокою називають різницю в заряді між внутрішньою та зовнішньою поверхнями плазмалеми[78]. Цьому сприяє різна проникність іонів через неї. Сама плазмалема пронизана іонними каналами — білковими «коридорами», через які іони вибірково здатні проходити[79]. В стані спокою у мембрані відкриті деякі калієві канали, тому К+ виходить зсередини клітини, де його більше, в позаклітинне середовище, де його менше. При цьому канали для позитивних іонів, які можуть увійти до клітини здебільшого закриті (хоча й не всі). На внутрішньому боці плазмалеми, незважаючи на високу кількість іонів калію, заряд негативний завдяки великим молекулам, які містять кислотні залишки та не можуть вийти з клітини крізь мембрану, та штучно підтримуваній заниженій концентрації протонів (pH цитоплазми слабколужний — 7,4). Негативний заряд на внутрішній стороні мембрани не дає К+ відходити далеко завдяки електростатичному тяжінню, що спричиняє надлишок позитивних йонів на зовнішній поверхні клітинної мембрани.[80][79] Вихід калію, однак, триває лише до того часу, коли встановлюється рівноважний електрохімічний потенціал (в контексті калію — калієвий потенціал), який обчислюється рівнянням Нернста. Калієвий потенціал — це стан, за якого дія електричного поля зрівноважує дифузійний тиск та унеможливлює рух калію (вихід калію спричиняє все більше зростання негативного заряду, що в певний момент унеможливлює вихід позитивних катіонів).[81][82]
Як вже було сказано, вхід іонів в клітину, хоч обмежений, але завдяки різниці концентрації відбувається. Найважливішим іоном, який прямує в клітину є Na+. Теоретично може наступити такий момент, коли різниця концентрацій іонів в клітині та поза нею зникне (через постійний вхід та вихід катіонів), і, як наслідок, зникне потенціал спокою, а потенціал дії неможливо буде згенерувати. Проте у здоровій клітині такого не відбувається, оскільки існує натрій-калієвий насос, який виводить з клітини три Na+ та приносить два K+ в клітину. Цей процес є енергозалежним (потребує розщеплення АТФ). Окрім іонів натрію та калію в генерації потенціалу спокою (і потенціалу дії) беруть участь, наприклад, Cl-, Ca2+, Mg2+. Окрім натрій-калієвого насосу існують інші іонні насоси.[81]
Потенціал дії
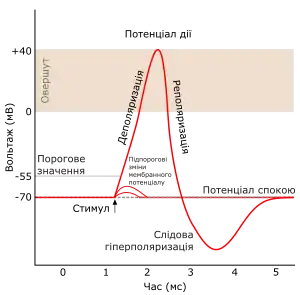
Потенціал дії — це короткочасна високоамплітудна деполяризація мембрани, тобто зменшення різниці потенціалів до нуля. Рух іонів призводить навіть до зміни знаку заряду на зовнішній (стає негативним) та внутрішній (стає позитивним) поверхні плазмалеми[83]. Потенціал дії нервових клітин часто називають нервовим імпульсом. З фізіологічної точки зору генерація потенціалу дії є збудженням, а здатність генерувати потенціали дії — збудливістю.
Появі потенціалу дії передує невеликий локальний вхід до клітини позитивних іонів — локальний деполяризаційний потенціал. Вхід Na+ і Ca+ відбувається через іонні канали, які відкриваються у відповідь на хімічні чи фізичні стимули. Коли різниця потенціалів зміниться до порогового значення (приблизно −50 мВ), яке дозволить відкритися потенціалзалежним натрієвим каналам, починається потенціал дії. Через ці канали в клітину спочатку потрапляє велика кількістю позитивних іонів — Na+, який рухається згідно з концентраційним градієнтом. Згодом відкриваються калієві канали аби відновити потенціал спокою, і з клітини виходять K+[84]. Розрізняють п'ять фаз потенціалу дії[83]: фаза деполяризації, фаза овершуту, фаза реполяризації, слідова реполяризація та слідова гіперполяризація. Виникнення потенціалу дії підпорядковується закону «все або нічого»: струм, сила якого має допорогове значення не здатна викликати потенціал дії («нічого»). У випадку дії струму, який перевищує пороговий рівень, виникає потенціал дії зі всіма стадіями («все»).[85]
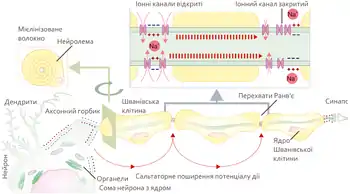
Поширення потенціалу дії по мієлінізованих та немієлінізованих волокнах буде різнитися. Так, у випадку немієлінізованого волокна імпульс буде поширюватися поступово, ділянка за ділянкою. Чим товстіше немієлінізоване волокно, тим швидше по ньому проходитиме імпульс.[86] У випадку мієлінізованого волокна потенціал дії поширюється лише від одного перехвату до іншого — стрибкоподібно, або сальтаторно, що значно пришвидшує поширення імпульсу.[87]
Рефлекси
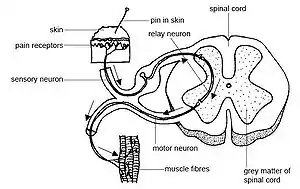
Рефлекс — це стереотипна реакція організму на зміни у зовнішньому або внутрішньому середовищах за участі нервової системи. Рефлекс є основою діяльності нервової системи.
Анатомічною основою рефлексу є рефлекторна дуга. Рефлекторна дуга — це шлях від рецептора до ефектора (органа-мішені, який має виконати певну дію). Отже, до складу рефлекторної дуги входять рецептори, аферентні волокна, нервовий центр (сукупність нейронів, які забезпечують виконання рефлексу), еферентні волокна та ефектор. В рефлекторних ланцюгах автономних рефлексів руховий нейрон міститься за межами ЦНС, у вегетативному вузлі[88]. В рецепторі виникає рецепторний потенціал, який здатний до сумації (є протилежністю потенціалу дії за своєю природою), який викликає потенціал дії в чутливому нейроні. Надалі потенціал дії прямує по ЦНС та ПНС завдяки синаптичній передачі. Через те, що передача в синапсах відбувається зі затримкою, час від моменту подразнення до моменту виникнення реакції називається латентним періодом.
Залежно від будови рефлекторної дуги рефлекси бувають моносинаптичними, коли дуга містить один чутливий та один руховий нейрони (відповідно наявний тільки один центральний синапс, а поняття «нервовий центр» є умовним), та полісинаптичними, коли центральних синапсів більше ніж один за рахунок наявності одного або декількох інтернейронів. Рефлекси також класифікують за органом-мішенню (рухові та вегетативні), за розміщенням рецепторів (інтеро-, екстеро- та пропріорецептивні рефлекси). Рефлекси бувають вродженими, або безумовними (наприклад, харчовий рефлекс) та набутими, або умовними — це рефлекси, які базуються на вроджених і виникають в процесі життєдіяльності. З клінічної точки зору рефлекси бувають нормальними та патологічними.[89]
Синапси
.jpg.webp)
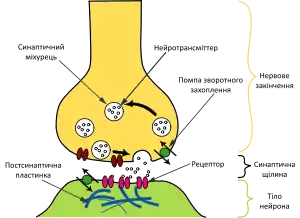
Місце передачі інформації від нейрона до нейрона чи від нейрона до м'язового волокна називається синапсом[90]. Основна кількість синапсів в нервовій системі тварин – це хімічні синапси[91]. Вони складаються з пре- та постсинаптичної мембран та синаптичної щілини[92]. В хімічному синапсі нервовий імпульс прямує тільки в одному напрямку: від пресинаптичної мембрани до постсинаптичної, завдяки виділенню нейромедіатора в синаптичну щілину[90][93].
Хімічні синапси бувають нервово-м'язовими та міжнейрональними[94]. Останні діляться на аксо-дендритні, аксо-соматичні, аксо-аксональні та дендрито-дендричні. Хімічні синапси також діляться в залежності від нейромедіатора, який використовується для передачі збудження.[92]
Нейромедіатори, або нейротрансмітери – це речовини, які забезпечують передачу сигналу від одного нейрона до іншого в хімічному синапсі. Вони бувають різної біохімічної структури (аміни, амінокислоти, нейропептиди).[92][95] За способом дії вони бувають іонотропними, тобто такими, які безпосередньо впливають на проникність іонних каналів та метаботропними – такими, які впливають на іонні канали через посередництво вторинних месенджерів. Перші викликають швидку синаптичну відповідь, а другі – повільну. Дію метаботропних нейромедіаторів також називають нейромодуляторною дією, оскільки вони змінюють процеси протікання синаптичної передачі (сприяють або запобігають виникненню потенціалу дії).[96][97]
Окрім хімічних синапсів існують електричні. В електричному синапсі між двома нейронами існує спеціальний білковий «коридор», через який іони (а отже і струм) та молекули без жодних перешкод прямують в обидві сторони.[91] В деяких тварин вивчені змішані синапси, в яких наявна як пряма електрична, так і хімічна передача імпульсу. Відмінною від синаптичної передачі (пряма взаємодія нейронів) є ефаптична (непряма взаємодія нейронів), яка реалізується завдяки близькому розташуванню нервових волокон та дії локальних струмів).
Сенсорна система
У процесі існування організм живої істоти постійно стикається з подразниками. Аналіз та відповідь на них є необхідною умовою для виживання. Системи, які це забезпечують (забезпечують відчуття) називаються сенсорними. Анатомічною основою сенсорних систем є рецептори (сприймальна частина), нервові шляхи (провідна частина) та мозкового центру (підкіркових та кіркових ділянок в головному мозку). Разом всі ці структури утворюють аналізатор. Найкраще рецептор реагує на адекватний подразник (палички та колбочки — на кванти світла, механорецептори — на дотик) — це називається специфічністю рецептора, хоча його потенціал може бути викликаний і неадекватним подразником (наприклад, сильний удар в око спричиняє «іскри»).
Хребетні, соматосенсорні системи яких дослідженні найкраще, здатні до таких відчуттів: зору, нюху, слуху, соматосенсорних (дотик, біль, температура), пропріорцепепторних (відчуття положення тіла) та інтерорецепторних (сигнали від внутрішніх органів).
Вивчення нервової системи
Нейронаука
Нейронаука складається з великої кількості дисциплін, які вивчають нервову систему з метою зрозуміти її будову, організацію, розвиток, зрозуміти, як в нервовій системі виникає свідомість, поведінкові явища. Так, нейроанатомія вивчає будову нервової системи, нейрохімія — хімічний склад нервової системи, нейрофізіологія — принципи функціонування нервової системи, порівняльна нейроанатомія — основні тенденції розвитку нервової системи в часовому вимірі, неврологія вивчає патологію нервової системи. Нейронаука не є суто біологічною наукою, оскільки взаємодіє з математикою, фізикою, філософією, соціологією та іншими небіологічними науками.[98]
Давній світ

Думки щодо функції, будови та важливості нервової системи змінювалися протягом давніх часів. Ще в часи неоліту проводилися трепанації[99]. Найвірогідніше, хірурги таким чином намагалися виганяти злих духів, які оволоділи людиною[100]. Цими «злими духами» могли бути напади епілепсії[101].
Вперше слово «мозок» згадується в папірусі Едвіна Сміта[102]. Проте у Стародавньому Єгипті мозок не вважали дуже важливим органом чи «центром», в якому містилася душа або щось подібне на неї. Про це свідчить той факт, що під час муміфікації мозок витягували через ніздрі та викидували геть.[103] Такі переконання довгий час панували і у Стародавній Греції. Головним органом вважалося серце, а більшість міфічних сил (noos, psyche та thymos) були розміщені в грудній клітці.[104] Першим, хто вказував на головний мозок, як центр свідомості, був Алкмеон Кротонський. Він також описав ряд нервів, серед яких зорові, та пов'язував мозок з відчуттями (однак нерви він називав poroi – каналами, в яких мала б бути порожнина).[105] Таке твердження щодо функції мозку підтримував Гіппократ. Він також надав анатомічний опис головного мозку, проте уявлення про роботу головного мозку в нього, як такі були відсутні. Він вважав, що кров насичена специфічною «життєвою силою», яка спричиняє рух та забезпечує розумну поведінку. Такі твердження існували через те, що уявлення про нервову систему, як таку, були відсутні. Послідовники Гіппократа описували нерви як poroi (канал) або tonoi (струна) і вважали, що за функцією вони подібні до вен.[106] Роль мозку обговорювали не тільки медики: Платон був прихильником того, що емоції та розум пов'язані з головним мозком[107], в той час як Арістотель відкидав такі ідеї та вважав, що серце є центром емоцій та інтелекту, який продукує тепло, а мозок служить лише для охолодження тіла[108]. Арістотель також був видатним зоологом і під час своїх дослідів на тваринах описав багато анатомічних структур головного мозку (півкулі, шлуночки, мозочок, мозкові оболони)[108].
Прорив у античних дослідженнях нервової системи відбувся в Александрії за часів Птолемеїв. Цей прорив пов'язаний з двома вченими – Герофілом та Ерасистратом, а також з тим фактом, що птолемейська Александрія була єдиним місцем в античному світі, де були дозволені розтини людських тіл.[109] Герофіл детально описав синуси твердої облони та шлуночки[110]. Проте найбільше його досягнення – це опис нервової системи, як такої. Він описав нерви, як ниткоподібні утворення, які відходять не від серця, а від мозку та спинного мозку, та назвав їх neura. Згідно з Галеном, він описав сім пар черепних нервів та розрізняв серед них рухові та чутливі.[111] Він також вважав, що життєва енергія міститься в шлуночках мозку[112]. Ерасистрат ще детальніше описав шлуночки мозку та стверджував про існування різних видів пневми; від нервів органи діставали фізичну пневму, яка забезпечувала рух та відчуття. Така пневма утворювалася в мозку.[113]
Кульмінацією античних досліджень стали праці Клавдія Галена. Він надав опис різних анатомічних структур (наприклад, гіпофіз), проте часто користувався працями інших авторів, оскільки досліди та розтини проводив на тваринах (так, надана ним класифікація черепних нервів була запозичена у Герофіла та Марінуса Александрійського).[114] Проте головне його досягнення – це досліди на тваринах. Так, при перетині нерва у свині він відкрив іннервацію голосових зв'язок[115], перерізав спинний мозок на різних рівнях та спостерігав за симптомами, які виникали[116], всіляко пошкоджував головний мозок[117]. Він також був прихильником теорії пневми, проте, на відміну від Ерасистрата, вважав, що фізична пневма утворюється в судинному сплетенні основи мозку і лише потім потрапляє в шлуночки[118]. Праці Галена мали великий вплив на медичну науку Середньовіччя і багато в чому були беззаперечними[119].
Примітки
- Foundational Model of Anatomy
- Price, 2011, с. 21.
- Price, 2011, с. 24.
- Gilbert, 2009, с. 333-334.
- Squire, 2008, с. 297.
- Gilbert, 2009, с. 335-336.
- Gilbert, 2009, с. 339.
- Gilbert, 2009, с. 373.
- Gilbert, 2009, с. 389.
- Price, 2011, с. 38-39.
- Gilbert, 2009, с. 374.
- Price, 2011, с. 36-37.
- Gilbert, 2009, с. 340-341.
- Головацький, 2 том, 2007, с. 212.
- Price, 2011, с. 31.
- Головацький, 2 том, 2007, с. 204.
- Squire, 2008, с. 42.
- Brodal, 2010, с. 5.
- Brodal, 2010, с. 6.
- Філімонов, 2010, с. 89-90.
- Squire, 2008, с. 18.
- Squire, 2008, с. 21.
- Verkhratsky, 2013, с. 74.
- Brodal, 2010, с. 19.
- Verkhratsky, 2013, с. 76.
- Brodal, 2010, с. 26.
- Verkhratsky, 2013, с. 79.
- Verkhratsky, 2013, с. 81.
- Головацький, 2 том, 2007, с. 203.
- Brodal, 2010, с. 411.
- Філімонов, 2010, с. 137.
- Головацький, 2 том, 2007, с. 238.
- Головацький, 3 том, 2009, с. 7.
- Маруненко, 2013, с. 19.
- Galizia, 2013, с. 179.
- Juergen K. Mai, George Paxinos. The Human Nervous System. — 3rd. — London : Elsiever Inc, 2011. — С. 143-144. — ISBN 978-0123742360.
- Філімонов, 2010, с. 141.
- Galizia, 2013, с. 180-181.
- Brodal, 2010, с. 413.
- Galizia, 2013, с. 183.
- Galizia, 2013, с. 197.
- Galizia, 2013, с. 200-202.
- Galizia, 2013, с. 205.
- Маруненко, 2013, с. 17.
- Squire, 2008, с. 16.
- Brusca, 2003, с. 250.
- Kaas, 2006, с. 300.
- Brusca, 2003, с. 279.
- Kaas, 2006, с. 301.
- Маруненко, 2013, с. 18.
- Brusca, 2003, с. 251.
- Brusca, 2003, с. 253.
- Richard A. Satterlie (2011). Do jellyfish have central nervous systems?. Journal of Experimental Biology 214 (8): 1215–1223. PMID 21430196. doi:10.1242/jeb.043687.(англ.)
- Squire, 2008, с. 17.
- Kaas, 2006, с. 302.
- Brusca, 2003, с. 300.
- Simon A. Levin. Encyclopedia of Biodiversity. — New York : Academic Press, 2013. — С. 460. — ISBN 978-0123847195. (англ.)
- Brusca, 2003, с. 302.
- Squire, 2008, с. 19-20.
- Verkhratsky, 2013, с. 79-80.
- Lewbart, 2011, с. 342.
- Маруненко, 2013, с. 20.
- Lewbart, 2011, с. 154.
- Brusca, 2003, с. 420.
- Brusca, 2003, с. 424.
- Brusca, 2003, с. 425.
- Lewbart, 2011, с. 161.
- Brusca, 2003, с. 492.
- Brusca, 2003, с. 570.
- Lewbart, 2011, с. 289-290.
- Kaas, 2006, с. 338.
- Brusca, 2003, с. 680.
- Brusca, 2003, с. 696.
- Brusca, 2003, с. 646.
- Маруненко, 2013, с. 20-21.
- Brusca, 2003, с. 619.
- Brusca, 2003, с. 571.
- Філімонов, 2010, с. 52.
- Squire, 2008, с. 112.
- Філімонов, 2010, с. 53.
- Філімонов, 2010, с. 54.
- Squire, 2008, с. 114.
- Філімонов, 2010, с. 55.
- Філімонов, 2010, с. 58.
- Шевчук, 2012, с. 34-35.
- Філімонов, 2010, с. 59.
- Філімонов, 2010, с. 60-61.
- Шевчук, 2012, с. 125.
- Шевчук, 2012, с. 53-55.
- Squire, 2008, с. 157.
- Brodal, 2010, с. 40.
- Шевчук, 2012, с. 56.
- Шевчук, 2012, с. 62-63.
- Squire, 2008, с. 228.
- Squire, 2008, с. 136.
- Brodal, 2010, с. 42-43.
- Squire, 2008, с. 227.
- Squire, 2008, с. 3.
- Wickens, 2014, с. 4.
- Wickens, 2014, с. 6.
- Wickens, 2014, с. 7.
- Wickens, 2014, с. 10.
- Wickens, 2014, с. 9.
- Wickens, 2014, с. 11.
- Wickens, 2014, с. 13.
- Wickens, 2014, с. 15.
- Wickens, 2014, с. 18.
- Wickens, 2014, с. 21-22.
- Wickens, 2014, с. 27-28.
- Wickens, 2014, с. 30.
- Wickens, 2014, с. 31.
- Wickens, 2014, с. 32.
- Wickens, 2014, с. 33.
- Wickens, 2014, с. 37-38.
- Wickens, 2014, с. 39-40.
- Wickens, 2014, с. 41.
- Wickens, 2014, с. 44.
- Wickens, 2014, с. 42-43.
- Wickens, 2014, с. 45.
Джерела
Українські
- Анатомія та еволюція нервової системи : підруч. [для студ. біол., філософ. (каф. психології) та мед. ф-тів ун-тів, пед. вузів] / Н. В. Федірко ; М-во освіти і науки, молоді та спорту України, Львів. нац. ун-т ім. І. Франка. – Л. : Вид-во ЛНУ, 2013. – 384 с. : іл. – (Серія "Біологічні Студії"). – Бібліогр.: с. 379-383. – ISBN 978-617-10-0011-7
- Головацький А. С., Черкасов В. Г., Сапін М. Р., Парахін А. І. Анатомія людини у трьох томах. — Вінниця : Нова Книга, 2007. — Т. 2. — 456 с. — 4000 прим. — ISBN 978-966-382-062-0.
- Людина : Навч. посібник з анатомії та фізіології / Львів. 2002. — 240 с.
- Головацький А. С., Черкасов В. Г., Сапін М. Р., Парахін А. І. Анатомія людини у трьох томах. — Вінниця : Нова Книга, 2009. — Т. 3. — 376 с. — 4000 прим. прим. — ISBN 978-966-382-181-8.
- І. М. Маруненко, Неведомська Є. О., Волковська Г. І. Анатомія, фізіологія, еволюція нервової системи. — Київ : Центр учбової літератури, 2013. — 184 с. — ISBN 978-617-673-136-8.
- Філімонов В. І. Фізіологія людини. — Київ : ВСВ «Медицина», 2010. — 776 с. — ISBN 978-617-505-070-5.
- В. Г. Шевчук, В. М. Мороз, С. М. Бєлан, М. Р. Гжегоцький, М. В. Йолтухівський. Фізіологія. — Вінниця : Нова Книга, 2012. — 448 с. — ISBN 978-966-382-375-1.
Іноземні
- Alexei Verkhratsky, Arthur Morgan Butt. Glial Physiology and Pathophysiology. — London : Wiley-Blackwell, 2013. — 560 с. — ISBN 978-0470978535. (англ.)
- Larry Squire, Darwin Berg, Floyd Bloom, Sascha du Lac. Fundamental Neuroscience. — 3rd. — London : Academic Press, 2008. — 1280 с. — ISBN 978-0123740199. (англ.)
- Giovanni Galizia, Pierre-Marie Lledo. Neurosciences - From Molecule to Behavior: a University Textbook. — Berlin : Springer Spektrum, 2013. — 736 с. — ISBN 978-3642107689. (англ.)
- Per Brodal. The Central Nervous System // — Oxford : Oxford University Press, 2010. — 608 с. — ISBN 978-0195381153. (англ.)
- Jon H. Kaas. Evolution of Nervous Systems, Four-Volume Set. — 3rd. — London : Academic Press, 2006. — 2000 с. — ISBN 978-0123925602. (англ.)
- Andrew P. Wickens. A History of the Brain: From Stone Age Surgery to Modern Neuroscience // — London : Psychology Press, 2014. — 404 с. — ISBN 978-1848723658. (англ.)
- Scott F. Gilbert. Developmental Biology. — 9th. — New York : Sinauer Associates, Inc, 2009. — 711 с. — ISBN 978-0878933846. (англ.)
- David Price, Andrew P. Jarman, John O. Mason, Peter C. Kind. Building Brains: An Introduction to Neural Development. — New York : Wiley, 2011. — 348 с. — ISBN 978-0470712290. (англ.)
- Gregory A. Lewbart. Invertebrate Medicine. — 2nd. — New York : Wiley-Blackwell, 2011. — 504 с. — ISBN 978-0813817583. (англ.)
- Richard C. Brusca, Gary J. Brusca. Invertebrates. — 2nd. — Sunderland : Sinauer Associates, 2003. — 936 с. — ISBN 978-0878930975. (англ.)
Посилання
- НЕРВОВА СИСТЕМА
- Нервная система//Большая советская энциклопедия. — 1969–1978
- The Human Brain Project Homepage(англ.)
- Kimball's Biology Pages, CNS(англ.)
- Kimball's Biology Pages, PNS(англ.)