Гістонацетилтрансферази
Гістонові ацетилтрансферази(Histone acetyltransferases (HATs)) - ферменти, що ацетилюють лізинові залишки на гістонах. ДНК обмотана навколо гістонів, тому, перенесення ацетильних груп на гістони може вимикати чи вмикати певні гени. Ацетилювання гістонів переважно підвищує експресію гену. Такий механізм зміни експресії генів є епігенетичним.
 GCN5 histone acetyltransferase domain homo24-mer, Human | |
| Ідентифікатори | |
|---|---|
| Код КФ | 2.3.1.48 |
| Номер CAS | 9054-51-7 |
| Бази ферментів | |
| IntEnz | IntEnz view |
| BRENDA | BRENDA entry |
| ExPASy | NiceZyme view |
| MetaCyc | metabolic pathway |
| KEGG | KEGG entry |
| PRIAM | profile |
| PDB structures | RCSB PDB PDBe PDBj PDBsum |
| Gene Ontology | AmiGO • EGO |
| Пошук | |
| PMC | статті |
| PubMed | статті |
| NCBI | NCBI proteins |
| CAS | 9054-51-7 |
Ацетилювання гістонів пов'язане з активацією транскрипції і асоційоване з еухроматином. На еухроматині, що є менш компактним за гетерохроматин, може відбуватись транскрипція за рахунок меншої щільності. Спершу вважали, що ацетилювання гістонів нівелює їх позитивний заряд, таким чином, зменшуючи спорідненість до ДНК. Але сучасні дослідження показують, що ацетилювання створює сайти для білок-білкових взаємодій. Гістонові ацетилтрансферази можуть ацетилювати не тільки гістони, а ще, наприклад, деякі ядерні рецептори та інші транскрипційні фактори.
Родини ГАТ
Гістонові ацетилтрансферази(ГАТ) традиційно поділяють на два класи на основі субклітинної локалізації. ГАТ А-типу розташовані в ядрі і залучені в регуляцію генної експресії шляхом ацетилювання нуклеосомних гістонів в хроматині. Вони містять бромодомен, який допомагає їм зв'язуватись з гістонами. Gcn5, p300/CBP і TAFII250 - приклади ГАТ А-типу. ГАТ В-типу розташовані в цитоплазмі та відповідальні за ацетилювання щойносинтезованих гістонів до їх асоціації в нуклеосоми.
Gcn5-споріднені N-ацетилтрансферази (GNATs)
ГАТи можна згрупувати в декілька різних родин, спираючись на гомологію послідовностей, структурні особливості та функції. Gcn5-споріднені N-ацетилтрансферази характеризуються наявністю бромодомену, вони ацетилюють лізини на гістонах Н2В, Н3 та Н4. Всі білки цієї родини мають чотири консервативні мотиви(A-D) в каталітичному домені. Крім каталітичного, ферменти цієї родини мають ще N-кінцевий домен, Ada2 домен та C-кінцевий бромодомен.
MYST
Родина MYST названа по першим буквам чотирьох її членів: MOZ, Ybf2 (Sas3), Sas2 і Tip60. Для ферментів даної родини характерна наявність цинкових пальців та хромодомену, вони ацетилюють гістони Н2А, Н3 та Н4. MOZ(monocytic leukemia zinc finger protein) виявляє онкогенні властивості у людей.
Інші
Крім попередніх родин гістонових ацетилтрансфераз, є кілька інших ферментів, що знайдені у вищих еукаріотів. До них належать p300/CBP, коактиватор ядерних рецепторів (e.g., ACTR/SRC-1), TAFII250, TFIIIC, Rtt109, та CLOCK. p300/CBP має кілька регіонів з цинковими пальцями та бромодомен і кількома висококонсервативними доменами схожий на GNATs, функціонує, як активатор транскрипції.
Коактиватори ядерних рецепторів
Три важливих коактиватори ядерних рецепторів - SRC-1, ACTR та TIF-2. SRC-1 (steroid receptor coactivator-1) може взаємодіяти з p300/CBP та PCAF, ацетилтрансферазний домен розташований в С-кінцевому регіоні.
Таблиця підумовує різницю між ферментами, експресію в різних організмах та структурні особливості.
| Родина | Організм | Асоційовані комплекси | Субстратна специфічність | Структурні особливості |
|---|---|---|---|---|
| GNAT | ||||
| Gcn5 | S. cerevisiae | SAGA, SLIK (SALSA), ADA, HAT-A2 | H2B, H3, (H4) | Bromodomain |
| GCN5 | D. melanogaster | SAGA, ATAC | H3, H4 | Bromodomain |
| GCN5 | H. sapiens | STAGA, TFTC | H3, (H4, H2B) | Bromodomain |
| PCAF | H. sapiens | PCAF | H3, H4 | Bromodomain |
| Hat1 | S. cerevisiae - H. sapiens | HAT-B, NuB4, HAT-A3 | H4, (H2A) | |
| Elp3 | S. cerevisiae | Elongator | H3, H4, (H2A, H2B) | |
| Hpa2 | S. cerevisiae | HAT-B | H3, H4 | |
| Hpa3 | S. cerevisiae | H3, H4 | ||
| ATF-2 | S. cerevisiae - H. sapiens | H2B, H4 | ||
| Nut1 | S. cerevisiae | Mediator | H3, H4 | |
| MYST | ||||
| Esa1 | S. cerevisiae | NuA4, piccolo NuA4 | H2A, H4, (H2B, H3) | Chromodomain |
| Sas2 | S. cerevisiae | SAS, NuA4 | H4, (H2A, H3) | |
| Sas3 (Ybf2) | S. cerevisiae | NuA3 | H3, (H4, H2A) | |
| Tip60 | H. sapiens | Tip60, NuA4 | H2A, H4, (H3) | Chromodomain |
| MOF | D. melanogaster | MSL | H4, (H2A, H3) | Chromodomain |
| MOZ | H. sapiens | MSL | H3, H4 | |
| MORF | H. sapiens | MSL | H3, H4 | |
| HBO1 | H. sapiens | ORC | H3, H4 | |
| p300/CBP | ||||
| p300 | H. sapiens | H2A, H2B, H3, H4 | Bromodomain | |
| CBP | H. sapiens | H2A, H2B, H3, H4 | Bromodomain | |
| SRC (nuclear receptor coactivators) | ||||
| SRC-1 | H. sapiens | ACTR/SRC-1 | H3, H4 | |
| ACTR (RAC3, AIB1, TRAM-1) | H. sapiens | ACTR/SRC-1 | H3, H4 | |
| TIF-2 (GRIP1) | H. sapiens | H3, H4 | ||
| SRC-3 | H. sapiens | |||
| Other | ||||
| TAFII250 (TAF1) | S. cerevisiae - H. sapiens | TFIID | H3, H4, (H2A) | Bromodomain |
| TFIIIC (p220, p110, p90) | H. sapiens | TFIIIC | H2A, H3, H4 | |
| Rtt109 | S. cerevisiae | Histone chaperones | H3 | |
| CLOCK | H. sapiens | H3, H4 |
Загальна структура
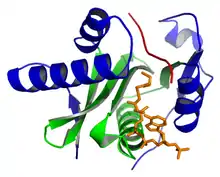
В загальному, ГАТи характеризуються структурно консервативним коровим регіоном, який утворюють 3 бета-складки, після яких розташована довга альфа-спіраль, повернута одним боком до бета-регіону. Основна область, яка відповідає мотивам A, B і D білків GNAT, фланкується на протилежних сторонах N- і C-кінцевими α / β сегментами, які є структурно унікальними для даної родини ГAT. Центральне ядро і бічні сегменти разом утворюють щілину над першим, де субстрати гістонів можуть зв'язуватися перед каталізом. У той час як домен центрального ядра (мотив A в GNATs) бере участь в зв'язуванні ацетил-CoA і каталізі, N- і C-кінцеві сегменти допомагають у зв'язуванні субстратів гістонів. Унікальні особливості, пов'язані з послідовністю та / або структурою N- і С-кінцевих областей для різних HAT-сімей, можуть допомогти пояснити деякі спостережувані відмінності серед HATs в специфічності субстрату гістонів. Припускається, що зв'язування CoA розширює паз для зв'язування гістонів в центральному ядрі шляхом переміщення C-кінцевого сегмента Gcn5 назовні. Крім того, оскільки контакти між CoA і білком сприяють утворенню сприятливих контактів гістонового білка, ймовірно, що зв'язування CoA передує зв'язуванню гістонів in vivo.
Гени деяких ГАТ
- KAT2A - кодує GCN5. Розташований на довгому плечі 17-ї хромосоми, складається з 8 286 нуклеотидів. Продукт - GCN5 складається з 837 амінокислот і має молекулярну масу в 93,9 кДа
- KAT2B - кодує PCAF. Розташований на короткому плечі 3-ї хромосоми, складається з 114 382 нуклеотидів. Продукт - PCAF складається з 832 амінокислот і має молекулярну масу в 93 кДа
- HAT1 - кодує HAT1. Розташований на довгому плечі 2-ї хромосоми, складається з 61 262 нуклеотидів. Продукт - HAT1 складається з 419 амінокислот і має молекулярну масу в 49,5 кДа
- KAT6A - кодує MOZ. Розташований на короткому плечі 8-ї хромосоми, складається з 122 512 нуклеотидів. Продукт - MOZ складається з 2004 амінокислот і має молекулярну масу в 225 кДа
- NCOA1 - кодує SRC-1. Розташований на короткому плечі 2-ї хромосоми, складається з 278 789 нуклеотидів. Продукт - SRC-1 складається з 1441 амінокислот і має молекулярну масу в 156,8 кДа
Біологічна роль
Ремоделювання хроматину
Гістонові ацетилтрансферази виконують багато біологічних функцій всередині клітини. Хроматин є комбінацією білків і ДНК, що знаходяться в ядрі, і він зазнає багатьох структурних змін, оскільки відбуваються різні клітинні події, такі як реплікація ДНК, репарація ДНК і транскрипція. Хроматин в клітині можна знайти в двох станах: конденсованому і неконденсованому. Останній, відомий як еухроматин, є транскрипційно активним, тоді як перший, відомий як гетерохроматин, є транскрипційно неактивним. Гістони являють собою білкову частину хроматину. Існує п'ять різних гістонових білків: H1, H2A, H2B, H3 і H4. Гістонові ядра утворюються, коли по два кожного підтипу гістонів, за винятком H1, утворюють четвертинний комплекс. Цей октамерний комплекс, разом з 147 парами основ ДНК, згорнутою навколо нього, утворює нуклеосому. Гістон H1 скріплює комплекс нуклеосом, і він є останнім білком, який зв'язується з комплексом..
Гістони, як правило, є позитивно зарядженими білками з N-кінцевими хвостами, які виходять з кору. Фосфодіефірний остов ДНК є негативним, що дозволяє проводити сильні іонні взаємодії між білками гістонів і ДНК. Ацетилтрансферази гістонів переносять ацетильну групу до специфічних залишків лізину на гістонах, що нейтралізує їх позитивний заряд і таким чином зменшує сильні взаємодії між гістонами і ДНК. Вважається також, що ацетилювання викликає збурення взаємодій між окремими нуклеосомами і діє як сайти взаємодії для інших пов'язаних з ДНК білків.
Можуть бути різні рівні ацетилювання гістонів, а також інші типи модифікацій, що дозволяють клітині контролювати рівень упаковки хроматину під час різних клітинних подій, таких як реплікація, транскрипція, рекомбінація та репарація. Ацетилювання не є єдиною регуляторною посттрансляційної модифікацією гістонів, що визначає структуру хроматину; також існують процеси метилювання, фосфорилювання, ADP-рибозилювання та убіквітинування. Ці комбінації різних ковалентних модифікацій на N-кінцевих хвостах гістонів називаються гістоновим кодом, і вважається, що цей код може бути спадковим і збережений в наступній генерації клітин.
Гістонові білки H3 і H4 є головними мішенями ГAT, але H2A і H2B також ацетильовані in vivo. Лізини 9, 14, 18 і 23 Н3 і лізинів 5, 8, 12 і 16 Н4 є орієнтованими на ацетилювання. Лізини 5, 12, 15 і 20 ацетилюються на Н2В, тоді як тільки лізини 5 і 9 виявляються ацетильованими на гістоні Н2А.
Експресія генів
Гістонові модифікації модулюють упаковку хроматину. Рівень упаковки ДНК важливий для транскрипції генів, оскільки доступ до промотору є важливим для того, щоб відбулася транскрипція. Нейтралізація заряджених залишків лізину ГAT дозволяє модифікувати хроматин таким чином, щоб цей РНК-полімераза мала доступ до транскрибованого гена. Однак ацетилювання не завжди пов'язане з посиленою транскрипционной активністю. Наприклад, ацетилювання H4K12 пов'язано з конденсованим і транскрипційно неактивним хроматином. Крім того, деякі модифікації гістонів пов'язані як з підвищеною, так і з репресованою активністю, залежно від умов.
ГАТи діють як транскрипційні коактиватори або генні інгібітори і найчастіше зустрічаються у великих комплексах. Ці комплекси включають SAGA (Spt / Ada / Gcn5L ацетилтрансфераза), PCAF, ADA (транскрипційний адаптер), TFIID (фактор транскрипції II D), TFTC (TAF комплекс без TBF) і NuA3 / NuA4 (нуклеосомні ацетилтрансферази Н3 і H4). Ці комплекси модулюють специфічність ГAT шляхом приведення ГAT до своїх цільових генів, де вони потім можуть ацетилювати нуклеосомні гістони. Деякі ГAT-транскрипційні ко-активатори містять бромодомени - 110-амінокислотний модуль, який розпізнає ацетильовані залишки лізину і функціонально пов'язаний з коактиваторами в регуляції транскрипції.
Див. також
Література
- Marmorstein R (August 2001). "Structure of histone acetyltransferases". Journal of Molecular Biology. 311 (3): 433–44. doi:10.1006/jmbi.2001.4859. PMID 11492997.
- Berger S.L. The complex language of chromatin regulation during transcription. Nature. 2007; 447: 407-412[1]
- https://doi.org/10.1016/j.bbagrm.2011.07.006
- https://doi.org/10.1016/j.prp.2017.06.013
- Su J, Wang F, Cai Y, Jin J. The Functional Analysis of Histone Acetyltransferase MOF in Tumorigenesis. Int J Mol Sci. 2016;17(1):99. Published 2016 Jan 14. doi:10.3390/ijms17010099
- Lee KK, Workman JL (April 2007). "Histone acetyltransferase complexes: one size doesn't fit all". Nature Reviews. Molecular Cell Biology. 8 (4): 284–95. doi:10.1038/nrm2145. PMID 17380162.
Посилання
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=KAT6A&keywords=moz
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=NCOA1&keywords=src-1
- https://www.genecards.org/cgi-bin/carddisp.pl?gene=KAT2B&keywords=pcaf
- https://www.genecards.org/Search/Keyword?queryString=pcaf
- Zhou, Dao-Xiu; Wei, Wei; Shen, Yuan (1 жовтня 2015). Histone Acetylation Enzymes Coordinate Metabolism and Gene Expression. Trends in Plant Science (English) 20 (10). с. 614–621. ISSN 1360-1385. PMID 26440431. doi:10.1016/j.tplants.2015.07.005. Процитовано 10 січня 2019.
