Еухроматин
Еухромати́н — стан хроматину (ДНК в ядрі) який характеризується "відкритістю" та транскрипційною активністю — з еухроматину зчитується РНК. В противагу еухроматину, гетерохроматин — стан ДНК, що щільно запакований і не є транскрипційно активним.
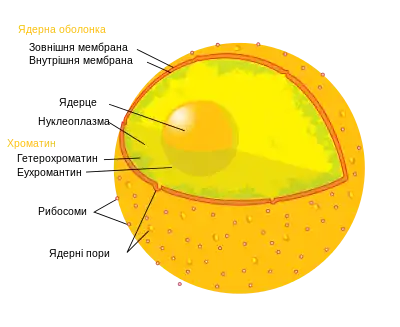
Склад еухроматину
Еухроматин, як будь-який хроматин, складається з білкового та ДНК компонентів.
ДНК
Особливістю ДНК, що входить до еухроматину, є те, що вона часто представлена у вигляді генів чи псевдогенів і не містить багато повторюваних ділянок, натомість складається з одиничних копій ДНК[1]. Також гетерохроматин в середньому містить більше метильованого цитозину, ніж еухроматин.
Метилювання ДНК у 5 положенні цитозину (5-метил-цитозин, 5mC) у послідовності нуклеотидів 5'-CG-3', що записується CpG, — одна з найвідоміших і добре вивчених епігенетичних модифікацій. Розподілення CpG послідовностей досить нерівномірне і трапляються їх скупчення в геномі, так звані CpG-острівці. При метилюванні CpG-острівців ДНК-метилтрансферазами спостерігяється встановлення гетерохроматин. Вилучення метильованого цитозину впродовж довгого часу вважалося лише пасивним — при реплікації ДНК у новосинтезованому ланцюзі ДНК вставляється неметильований цитозин. Проте з першим десятиріччям XXI сторіччя стає зрозуміло, що є недокінця встановлені механізми активного деметилювання ДНК. Для встановлення еухроматину послідовності ДНК повинні бути звільненими від 5mC[2]
Гістони
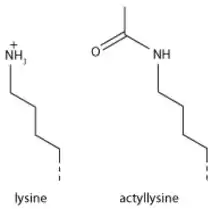
ДНК в ядрі не знаходиться у довільному стані, а накручується на нуклеосоми. Хромосома складається з великої кількості нуклеосом, на які накручена ДНК, а в залежності від того, наскільки щільно нуклеосоми прилягають одна до одної і формують додаткові, більш щільно запаковані структури, формується гетеро- чи еухроматин.
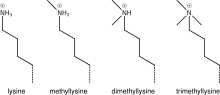
Характерною особливістю білкового складу еухроматину є посттрансляційні модифікації гістонів, які складають нуклеосоми, на яких накручується ДНК. Еухроматину притаманне одно-, дву- та триметилювання гістону H3 по 4 залишку лізину H3K4me, тоді як гетерохроматин триметельований по 9 залишку лізину (H3K9me)[3]. Також гістони H3 та H4 часто мають ацетильовані хвости, тоді як у гетерохроматиновій ділянці ці гістони часто гіпоацетильовані. На відміну від метилювання гістонів, що не змінює заряду амінокислотного залишку, ацетилювання залишку лізину прибирає позитивний заряд з амінокислоти. Це призводить до зниження електростатичного притягання негативнозарядженої молекули ДНК до вже електро-нейтральної амінокислоти. Таким чином молекулу ДНК легше від'єднати від нуклеосоми, на яку вона намотана і доступність до даної ділянки ДНК зростає[4]
Формування гетерохроматину часто починається на певній ділянці ДНК, і далі розповсюджуватися по хромосомі. Для запобігання перетворення гетерохроматину на еухроматин і еухроматину на гетерохроматин у клітин деяких організмів знайдені бар'єрні ДНК ділянки, які розмежовують гетеро- та еухроматинові частини хромосом. Так у курки спеціальний фактор транскрипції USF1 з'єднується з бар'єрною ділянкою на 5'-кінці від локусу β-globin, який оточений гетерохроматином, та призводить до залучення факторів, що запобігають конденсуванню хроматину у цьому місці. Ці фактори є білками гістонацетилтрансферази, гістонметилтрансферази що метилюють H3 по 4 лізину[3].
Проте не у всіх організмів є чіткі розмежування конденсованого і деконденсованого хроматину. У дрозофіли встановлення гетеро- або еухроматину вирішується місцево в залежності від локальної активності та концентрації ферментів, які конденсують чи деконденсують хроматинову фібрилу[3].
Порівняння еу- та гетерохроматину
| Пункт | Еухроматин | Гетерохроматин |
| ДНК | гени, псевдогени | Повторювальні ділянки ДНК: транспозони, сателітна ДНК, теломери[3] |
| Положення | Ділянки транскрипції, промотори, екзони та інтрони активних генів, енхансери[5]. | Теломери, при-центромерні ділянки, тільце Барра |
| Модифікації гістонів* | H3K4me, H4R3me[3], H3K4me2, H3K4me3, H2B-ubiq, H3K36me, H3K79me, H3K9ac[5] | H3K9me[3], H3K9me2, H3K9me3[5] |
| Білкові взаємодії | білки пов'язані з транскрипцією РНК, зв'язуванням з промоторами, транскрипційні фактори тощо | HP1, Polycomb-комплекс |
* В таблиці під модифікаціями гістонів слід читати запис H#X#yz, де:
- H# гістон з його порядковим номером: 2B, 3, 4 — гістони H2B, H3 та H4 відповідно;
- X — латинський символ який відповідає залишку амінокислоти (див. таблицю-розшифровку символів для кожної амінокислоти, пункт «Назва»), K — лізин, R — аргінін;
- # — порядковий номер залишку амінокислоти у поліпептидному ланцюзі гістону;
- yz — модифікація, що відбувається з гістоном: me — метилювання (додавання метильної групи), me2, me3 — ди- та три-метилюванння (додавання двох та трьох метильних груп, відповідно), ubiq — убіквітинування (додавання убіквітину), ac — ацетилування (додавання ацетильної групи)
Так запис H3K4me розшифровується, як метилювання 4го лізину H3 гістону, а запис H4R3me — метилювання 3го аргініну H4 гістону
Див. також
Примітки
- Irene Chiolo, Jonathan Tang, Walter Georgescu & Sylvain V. Costes (October 2013). Nuclear dynamics of radiation-induced foci in euchromatin and heterochromatin. Mutation research 750 (1-2): 56–66. PMID 23958412. doi:10.1016/j.mrfmmm.2013.08.001.
- Peter A. Jones (July 2012). Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nature reviews. Genetics 13 (7): 484–492. PMID 22641018. doi:10.1038/nrg3230.
- Hisashi Tamaru (July 2010). Confining euchromatin/heterochromatin territory: jumonji crosses the line. Genes & development 24 (14): 1465–1478. PMID 20634313. doi:10.1101/gad.1941010.
- Johannes Graff & Li-Huei Tsai (February 2013). Histone acetylation: molecular mnemonics on the chromatin. Nature reviews. Neuroscience 14 (2): 97–111. PMID 23324667. doi:10.1038/nrn3427.
- А. В. Сиволоб, К. С. Афанасьєва (2012). Молекулярна організація хромосом. К: Видавничо-поліграфічний центр "Київський університет". с. 41.
Джерела
- А. В. Сиволоб, С.Р. Рушковський, С.С. Кир'яченко та ін. (2008). Генетика. К: Видавничо-поліграфічний центр "Київський університет".
- А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр "Київський університет".
- А. В. Сиволоб, К. С. Афанасьєва (2012). Молекулярна організація хромосом. К: Видавничо-поліграфічний центр "Київський університет".