Деградація РНК
Деградація РНК — загалом означає процес розкладання полімеру рибонуклеїнової кислоти на складові мономери, якими є рибонуклеотидфосфати (NMPs), у вужчому сенсі — ензиматичне розщеплення матричної РНК (мРНК) за участі спеціалізованих білкових комплексів клітини, таких як екзосома, P-тільця та стресові гранули[1]. Згідно із положеннями центральної догми молекулярної біології, матрична РНК виконує ключову роль в процесі трансляції протеїнів. Отже тривалість перебування певної мРНК в цитоплазмі визначає кількість протеїну, який з неї може бути напрацьований. Відповідно, за допомогою комплексів, що здійснюють деградацію РНК, клітина має змогу контролювати рівень експресії білок-кодуючих генів. Така регуляція називається прост-транскрипційною, оскільки відбувається вже після синтезу ДНК-залежною РНК-полімеразою певної матричної РНК. Слід зазначити, що кожна мРНК характеризується певним часом життя (напіврозпаду) в цитоплазмі. Окрім регулювання рівнів продукування різних білків, системи деградації РНК також здійснюють контроль якості новоутворених мРНК[2] шляхом утилізації тих мРНК, які не пройшли процесинг (зокрема, не можуть бути правильно сплайсовані), мають небажані вторинні структури або неправильно зв'язуються із РНК-зв'язуючими протеїнами. Разом із тим в клітині існують спеціальні системи розпізнавання та знешкодження чужинної РНК, які працюють за принципом РНК інтерференції.
Основні шляхи деградації матричної РНК
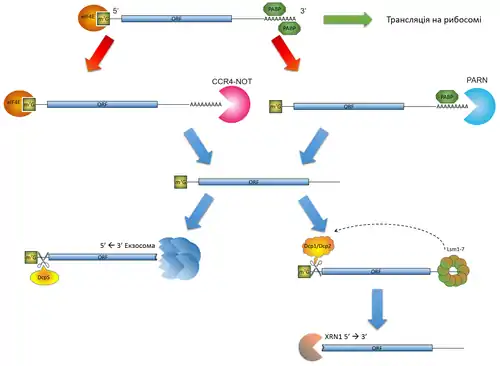
Кожна зріла матрична РНК, яка пройшла процесинг, має 5’-метилгуанозиновий кеп та 3'-поліаденозиновий хвіст. Ці структури зв'язуються зі спеціальними протеїнами: перша — із евкаріотичним фактором ініціації трансляції eIF4E[3], друга — із полі(А)-зв'язуючим протеїном PABP[4]. Додатково до згаданих факторів із мРНК можуть взаємодіяти інші РНК-зв'язуючі протеїни, які розпізнають послідовності на 3’- та 5’-нетрансльованих регіонах. Слід відмітити, що РНК в клітині ніколи не перебуває у вільному від білків стані, а натомість існує лише у формі рибонуклеопротеїнів[5]. У купі зазначені структури та білкові фактори захищають мРНК від передчасної деградації рибонуклеазами, а також допомагають взаємодіяти із рибосомою та сприяють трансляції.
Таким чином, для проходження процесу деградації, кожна мРНК має бути позбавлена захисних структур на 5’- та 3’-кінцях.
Відповідно, існує три основні шляхи деградації РНК[6]:
- видалення полі(А)-хвоста із наступним розщепленням в напрямку 3’ --> 5’,
- видалення 5’-метилгуанозинового кепу (декепінг) із наступним розщепленням в напрямку 5’ --> 3’
- ендонуклеазне розщеплення із подальшою деградацією за участі екзонуклеаз по новоутворених вільних 5’ та 3’ кінцях.
Кожен з цих механізмів забезпечується своїм набором ензимів, проте зазначені шляхи не є взаємовиключними і часто йдуть паралельно чи послідовно.
Деаденілювання
Основним шляхом деградації РНК в евкаріотів є видалення полі(А)-хвоста — деаденілювання[7]. За вкорочення полі(А)-тракту відповідає білковий комплекс CCR4-NOT або полі(А)-специфічна рибонуклеаза PARN. Причому, CCR4-NOT виконує деаденілювання лише за умови відсутності полі(А)-зв'язуючих протеїнів на хвості, тоді як PARN — лише у випадку наявності вільного 5’-метилгуанозинового кепу, не зв'язаного із eIF4E[6]. Обидві умови зазвичай виконуються для мРНК, яка щойно завеншила трансляцію. Слід відзначити, що деаденілювання — зворотний процес. Існують також цитоплазматичні полі(А)-полімерази, які здатні подовжувати полі(А)-хвіст, що дозволяє мРНК повернутися до трансляції.
Надалі деаденільована РНК може бути розщеплена двома шляхами: з 3’-кінця або з 5’-кінця. Перший передбачає розпад РНК у напрямку 3’ --> 5’ за участі комплексу екзонуклеаз під назвою екзосома. Під час 3’ --> 5’ деградації РНК в екзосомі 5’-метилгуанозиновий кеп від'єднується спеціальним кеп-скавенжером DcpS. Відсутність полі(А)-хвоста також може слугувати сигналом для ензимів, які напряму здійснюють декепінг (Dcp1 та Dcp2), що визначає другий шлях розпаду деаденільованої РНК. В такому разі спочатку відбувається розпізнавання деаденільованої РНК комплексом семи протеїнів Lsm1-7, котрий асоціюється із вільнім від полі(А) 3’-кінцем РНК. У подальшому Lsm1-7 залучає Dcp1 та Dcp2, які здійснюють видалення кеп-структури, а потім йде розщеплення РНК у напрямку 5’ --> 3’ за участі екзонуклеази XRN1[7].
Допоміжні шляхи деградації матричної РНК
Декепінг, незалежний від деаденілювання
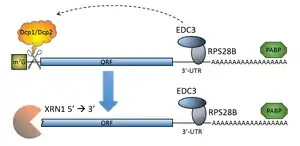
У дріжджів також відомий механізм розщеплення РНК, який по суті являє собою декепінг, але при цьому не залежить від деаденілювання. Натомість залучення декепінг-ензимів відбувається не з допомогою Lsm1-7 комплексу, а за участі білків, які розпізнають шпилькоподібну структуру на 3’-нетрансльованому регіоні мРНК. Такий механізм описаний для небагатьох мРНК Saccharomyces cerevisiae, наразі відомо щонайменше два транскрипти, що регулюються подібним чином, це — RPS28B та EDC1. Цікавим є той факт, що продукти обох цих генів самі безпосередньо беруть участь в процесі декепінгу, що незалежить від деаденілювання. Це може вказувати на авторегуляцію допоміжних шляхів деградації мРНК[8].
Розщеплення ендонуклеазами
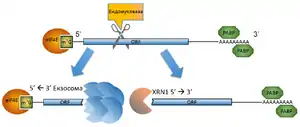
Ендонуклеолітичний розпад мРНК є найшвидшим і найефективнішим шляхом деградації, оскільки продуктом лише одного каталітичного акту ендонуклеази є відразу два вільні кінці — 3’ та 5’ —, які можуть надалі розщеплюватися екзосомою та XRN1 екзонуклеазою відповідно. Однак клітина вдається до нього здебільшого у надзвичайних випадках, як от різка перебудова метаболізму під час диференціювання або стресу.
Так наприклад ендонуклеаза PMR1 відповідає за розщеплення мРНК альбуміну у відповідь на стимуляцію естрогенового рецептору в ооцитах Xenopus laevis. В структурі PMR1 наявні домени зв'язування із полісомами, тож вважалося, що вона здатна розщеплювати мРНК під час трансляції. Але згодом було з'ясовано, що згадана ендонуклеаза також задіяна у формуванні стресових гранул — спеціальних компартментів у цитоплазмі, де відбувається затримка, сортування та диференційна деградація певних мРНК під час стресу[9].
Інша ендонуклеаза MRP, яка здебільшого задіяна в обробці рибосомальних та мітохондріальних РНК, також бере участь у деградації мРНК CLB2, що кодує циклін Б-типу, наприкінці мітозу. MRP розрізає CLB2 на ділянці 5’-нетрансльованого регіону, позбавляючи її таким чином кепу і викликаючи подальше розщеплення за допомогою екзонуклеази XRN1. Рецесивні мутації в гені ендонуклеази MRP описані у хворих на гіпоплазію хряща та волосся (cartilage-hair hypoplasia). Разом із тим у пацієнтів спостерігається підвищений рівень експресії цикліну CLB2. Отже порушення проліферації певних типів клітин відбувається через надмірну продукцію протеїну CLB2, що викликана відсутністю нормальної деградації відповідної мРНК у зв'язку із дефектом MRP ендонуклеази[10].
Див. також
Примітки
- Tourrière, Hélène; Chebli, Karim; Tazi, Jamal (2002). mRNA degradation machines in eukaryotic cells. Biochimie 84 (8): 821–837. ISSN 03009084. doi:10.1016/S0300-9084(02)01445-1.
- Vanacova, Stepanka; Stef, Richard (2007). The exosome and RNA quality control in the nucleus. EMBO reports 8 (7): 651–657. ISSN 1469-221X. doi:10.1038/sj.embor.7401005.
- von der Haar, Tobias; Gross, John D; Wagner, Gerhard; McCarthy, John E G (2004). The mRNA cap-binding protein eIF4E in post-transcriptional gene expression. Nature Structural & Molecular Biology 11 (6): 503–511. ISSN 1545-9993. doi:10.1038/nsmb779.
- Goss, Dixie J.; Kleiman, Frida Esther (2013). Poly(A) binding proteins: are they all created equal?. Wiley Interdisciplinary Reviews: RNA 4 (2): 167–179. ISSN 17577004. doi:10.1002/wrna.1151.
- Glisovic, Tina; Bachorik, Jennifer L.; Yong, Jeongsik; Dreyfuss, Gideon (2008). RNA-binding proteins and post-transcriptional gene regulation. FEBS Letters 582 (14): 1977–1986. ISSN 00145793. doi:10.1016/j.febslet.2008.03.004.
- Garneau, Nicole L.; Wilusz, Jeffrey; Wilusz, Carol J. (2007). The highways and byways of mRNA decay. Nature Reviews Molecular Cell Biology 8 (2): 113–126. ISSN 1471-0072. doi:10.1038/nrm2104.
- Norbury, Chris J. (2013). Cytoplasmic RNA: a case of the tail wagging the dog. Nature Reviews Molecular Cell Biology 14 (10): 643–653. ISSN 1471-0072. doi:10.1038/nrm3645.
- Badis, Gwenael; Saveanu, Cosmin; Fromont-Racine, Micheline; Jacquier, Alain (2004). Targeted mRNA Degradation by Deadenylation-Independent Decapping. Molecular Cell 15 (1): 5–15. ISSN 10972765. doi:10.1016/j.molcel.2004.06.028.
- Yang, F.; Peng, Y.; Murray, E. L.; Otsuka, Y.; Kedersha, N.; Schoenberg, D. R. (2006). Polysome-Bound Endonuclease PMR1 Is Targeted to Stress Granules via Stress-Specific Binding to TIA-1. Molecular and Cellular Biology 26 (23): 8803–8813. ISSN 0270-7306. doi:10.1128/MCB.00090-06.
- Tina Gill, Ti Cai, Jason Aulds, Sara Wierzbicki & Mark E. Schmitt (February 2004). RNase MRP cleaves the CLB2 mRNA to promote cell cycle progression: novel method of mRNA degradation. Molecular and cellular biology 24 (3): 945–953. PMID 14729943.