Матрична рибонуклеїнова кислота
Ма́трична рибонуклеї́нова кислота́ (мРНК, менше-вживаний синонім — інформаційна РНК або іРНК) — тип РНК, що відповідає за перенесення інформації про первинну структуру білків від ДНК до місць синтезу білків.
Загальний опис
мРНК синтезується на матриці ДНК в ході процесу транскрипції, після чого, у свою чергу, використовується в ході трансляціїї як матриця для синтезу білків. Тим самим мРНК грає важливу роль в експресії («прояві») генів.
Довжина типової зрілої мРНК складає від кількох сотень до кількох тисяч нуклеотидів. Щонайдовші мРНК відмічені у (+)оц РНК-вмісних вірусів, наприклад пікорнавірусів, проте слід пам'ятати, що у цих вірусів мРНК утворює весь їхній геном. ДНК нерідко порівнють з кресленнями для виготовлення білків. Розвиваючи цю інженерно-виробничу аналогію, можна сказати, що, якщо ДНК — це повний набір креслень для виготовлення білків, що знаходиться на зберіганні в сейфі директора заводу, то мРНК — тимчасова робоча копія креслення, що видається в складальний цех.
Гіпотеза про значення РНК в синтезі білків була висловлена Торбйорном Касперсоном (Torbjörn Caspersson) на основі досліджень 1937–1939 років, в результаті яких було показано, що клітини, що активно синтезують білки, містять велику кількість РНК. Підтвердження гіпотези було отримане Юбером Шантреном (Hubert Chantrenne).
«Життєвий цикл» мРНК
Життєвий цикл молекули мРНК починається транскрипцією і завершується деградацією. Молекула мРНК протягом свого життя може бути також оброблена і «відредагована» перед трансляцією. Еукаріотичні молекули мРНК часто вимагають складної обробки (процесингу) і транспортування, тоді як прокаріотичні молекули мРНК зазвичай цього не вимагають.
Транскрипція
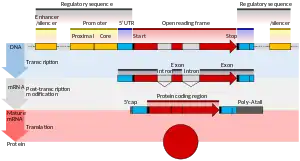
Транскрипцією називають процес копіювання генетичної інформації з ДНК на мРНК. Він здійснюється ферментом РНК-полімеразою, що «розплітає» подвійну спіраль ДНК і будує, згідно з принципом комплементарності, копію ділянки ДНК на основі одного з ланцюжків подвійної спіралі. Цей процес як у еукаріотів, так і у прокаріотів організований однаково. Основна відмінність між про- і еукаріотами полягає в тому, що у еукаріотів РНК-полімераза під час транскрипції асоціюється з мРНК-оброблювальними ферментами, тому у них процесинг мРНК і транскрипція можуть проходити одночасно. Короткоіснуючі необроблені або частково оброблені продукти транскрипції називаються передвісниками мРНК або пре-мРНК, після повного завершення процесингу — зрілою мРНК.
Процесинг еукаріотичної пре-мРНК
Тоді як мРНК прокаріотів (бактерій і архей), за рідкісними виключеннями, відразу готові до трансляції і більше не вимагають ніяких змін, еукаріотичні пре-мРНК вимагають значного процесингу. У процесі сплайсингу з пре-мРНК видаляються інтрони, на 5'-кінець додається кеп, на 3'-кінець додається поліаденіновий хвіст.
Сплайсинг
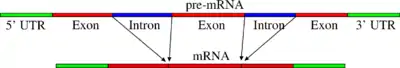
Сплайсинг — це процес, при якому з мРНК видалаються інтрони — послідовності нуклеотидів, що не кодують білок; залишаються лише екзони — ділянки послідовностей нуклеотидів ДНК, що кодують певний білок. Іноді пре-мРНК можуть бути сполучені різними способами, дозволяючи одному гену кодувати декілька білків. Цей процес називається альтернативним сплайсінгом. Сплайсінг зазвичай проводиться РНК-білковим комплексом, що називається сплайсосома, але деякі молекули мРНК також можуть каталізувати сплайсінг (див. рибозими).
Транспорт
Інша відмінність між еукаріотами і прокаріотами — транспорт мРНК. Через те, що еукаріотична транскрипція і трансляція просторово розділені, еукаріотичні мРНК повинні бути виведені з ядра в цитоплазму. Зрілі мРНК розпізнаються за наявністю модифікацій і покидають ядро через ядерні пори, в цитоплазмі мРНК утворюють нуклеопротеїдні комплекси — інформосоми, у складі яких транспортуються до рибосом.
Трансляція
Оскільки прокаріотичні мРНК не потребують процесингу і транспортування, трансляція рибосомою може початися негайно після транскрипції. Отже, трансляція у прокаріот у більшості випадків суміщена з транскрипцією і відбувається ко-транскрипційно.
Еукаріотична мРНК повинна бути оброблена і доставлена з ядра в цитоплазму, і лише тоді може бути трансльована рибосомою. Трансляція може відбуватися як на рибосомах, що знаходяться в цитоплазмі у вільному стані, так і на рибосомах, що асоціюються із стінками ендоплазматичного ретикулума. Таким чином, у еукаріотів трансляція ніколи не суміщена безпосередньо з транскрипцією.
Регуляція трансляції
Оскільки у прокаріотів транскрипція зазвичай суміщена з трансляцією, прокаріотична клітина може швидко реагувати на зміни в навколишньому середовищі шляхом синтезу нових білків, тобто регуляція відбувається, в основному, на рівні транскрипції. У еукаріотів через необхідність редагування і транспорту мРНК, відповідь на зовнішні стимули займає більше часу. Тому їхній синтез білків також інтенсивно регулюється і на пост-транскрипційному рівні. Не всяка зріла мРНК транслюється або транслюється на різному рівні, якраз через ці механізми регуляції експресії білків, наприклад РНК-інтерференція.
Деградація
Після деякого часу, що визначається її нуклеотидною послідовністю, зокрема, довжиною поліаденінової ділянки на 3'-кінці молекули, мРНК руйнується на нуклеотиди, з яких вона збудована, за участю РНКаз. Як правило, руйнування починається з видалення кепа на 5'-кінці, поліаденінового хвоста на 3'-кінці і потім екзонуклеазы одночасно руйнують мРНК в 5' -> 3' і 3' -> 5' напрямках. мРНК, в якій сигнал завершення синтезу білка, стоп-кодон, в результаті помилки транскрипції знаходиться в середині кодуючої послідовності, схильна до особливої швидкої форми деградації, НМД (nonsense-mediated decay), що проходить через зв'язування багатих на AU мотивів із специфічними білками[1].
Будова зрілої мРНК

Зріла мРНК складається з кількох ділянок, що розрізняються за функціями: «5' кеп», 5' нетрансльована область, кодуюча (трансльована) область, 3' нетрансльована область і 3' поліаденіновий «хвіст».
5' Кеп
5' кеп (або кап) (від англ. cap — шапочка) — це модифікований гуаніновий нуклеотид, який додається на 5' (передній) кінець незрілої мРНК. Ця модифікація дуже важлива для пізнавання мРНК при ініциації трансляції, а також для захисту від 5’-нуклеаз — ферментів, що руйнують ланцюжки нуклеїнових кислот з незахищеним 5'-кінцем.
Кодуючі області
Кодуючі області складаються з кодонів — послідовностей з трьох нуклеотідов, що слідують безпосередньо одна за одною, кожна з яких відповідає в генетичному коді певній амінокислоті або початку і кінцю синтезу білка. Кодуючі області починаються із старт-кодона і закінчуються одним з можливих стоп-кодонів. Зчитування послідовності кодонів і збірка на її основі послідовності амінокислот молекули білка, що синтезується, здійснюється рибосомами за участю транспортних РНК в процесі трансляції. На додаток до кодування білків, частини кодуючих областей можуть служити управляючими послідовностями. Наприклад, вторинна структура РНК в деяких випадках визначає результат трансляції.
Моноцистронна і поліцистронна мРНК
мРНК називають моноцистронною, якщо вона містить інформацію, необхідну для трансляції тільки одного білка (один цистрон). Поліцистронна мРНК кодує декілька білків. Гени (цистрони) в такій мРНК розділені інтергенними, некодуючими послідовностями. Поліцистронні мРНК характерні для прокаріотів і вірусів, у еукаріотів більша частина мРНК є моноцистронними[2].
Нетрансльовані області
Нетрансльовані області — ділянки РНК, розташовані до старт-кодона і після стоп-кодона, які не кодують білок. Вони називаються 5'-нетрансльованою областю і 3'-нетрансльованою областю, відповідно. Ці області транскрибуются у складі того ж самого транскрипту, що і кодуюча ділянка. Нетрансльовані області мають кілька функцій в життєвому циклі мРНК, включаючи регуляцію стабільності мРНК, локалізації мРНК і ефективності трансляції. Стабільність мРНК може контролюватися 5'- і/або 3'-область через різну чутливість до ферментів, які відповідають за деградацію РНК — РНКаз і регуляторних білків, які прискорюють або уповільнюють деградацію.
3' поліаденіновий хвіст
Довга (часто декілька сотень нуклеотидів) послідовність аденінових основ, присутня на 3' «хвості» мРНК еукаріотів, синтезується ферментом поліаденілат-полімеразою. У вищих еукаріотів полі-А-хвіст додається до транскрибованної РНК, яка містить специфічну послідовність, AAUAAA. Важливість цієї послідовності можна побачити на прикладі мутації в гені людського 2-глобіну, яка змінює AAUAAA на AAUAAG, що приводить до недостатньої кількості глобіну в організмі[3].
Вторинна структура
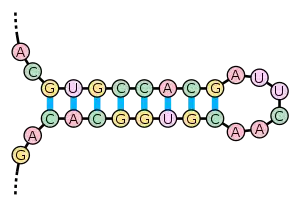
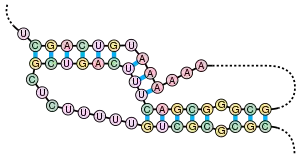
Окрім первинної структури (послідовності нуклеотидів), мРНК характеризується і вторинною структурою. На відміну від ДНК, вторинна структура якої заснована на міжмолекулярних взаємодіях (подвійна спіраль ДНК утворена двома лінійними молекулами, сполученими одна з одною на всій довжині водневими зв'язками), вторинна структура мРНК заснована на внутрішньомолекулярних взаємодіях (лінійна молекула «складається», і водневі зв'язки виникають між різними ділянками однієї і тієї ж молекули).
Прикладами вторинної структури можуть служити стебло-петля і псевдовузол.
Вторинні структури в мРНК служать для регуляції трансляції. Наприклад, вставка в білки нестандартних амінокислот, селенометіоніну і пірролізину залежить від стебла-петлі, розташованою в 3'-нетрансльованій області. Псевдовузли служать для програмованої зміни рамки зчитування генів.
У вірусних мРНК складні вторинні структури (IRES) направляють трансляцію, не залежну від розпізнавання кепа і факторів ініціації трансляції.
Див. також
- Деградація РНК
- Генетичний код
- Вірус
- Полісома
Література
- А. В. Сиволоб, С.Р. Рушковський, С.С. Кир'яченко та ін. (2008). Генетика. К: Видавничо-поліграфічний центр "Київський університет".
- А. В. Сиволоб (2008). Молекулярна біологія. К: Видавничо-поліграфічний центр "Київський університет".
- Козлов, Н. Н., Кугушев, Е. И., Сабитов, Д. И., Энеев, Т. М. (2002). Компьютерный анализ процессов структурообразования нуклеиновых кислот. Москва. (рос.)
Примітки
- A conserved AU sequence from the 3' untranslated region of GM-CSF mRNA mediates selective mRNA degradation. Cell 46 (5): 659–667. 1986. PMID 3488815. Текст «Shaw, G. and Kamen, R.» проігноровано (довідка)
- Kozak, M. (1983). Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles. Microbiological Reviews 47 (1): 1–45. PMID 6343825.
- Higgs DR, Goodbourn SE, Lamb J, Clegg JB, Weatherall DJ, Proudfoot NJ (1983). α-thalassaemia caused by a polyadenylation signal mutation. Nature 306 (5941): 398–400. PMID 6646217. doi:10.1038/306398a0.