Закони Менделя
Зако́ни Ме́нделя — закони, що є основою класичної генетики. У своїх працях Грегор Мендель ґрунтувався на дослідженнях, проведених на горосі посівному (рід Pisum). Цей об'єкт виявився вдалим, тому що для нього характерне самозапилення, яке уможливлює одержання чистих ліній, тобто особин гомозиготних за більшістю генів. У своїх роботах Мендель не виділяв окремих законів, їх виділили й назвали інші дослідники, вже після їхнього перевідкриття в 1900 році.

Передісторія
Уявлення про спадковість до робіт Менделя
Навіть у найдавніших писемних пам'ятках, що дійшли до наших днів, згадуються деякі особливості спадковості, такі як схожість між членами однієї сім'ї. Основні уявлення про спадковість, поширені до початку XX століття, базувались на двох принципах. По-перше, вважалось, що спадковість працює тільки в межах одного виду. Хоч у міфології багатьох народів стрічаються такі гібридні тварини як мінотаври, кентаври, мантикори тощо, а жирафи вважались наслідком схрещення верблюда і леопарда (це відображено у видовій назві Giraffa camelopardalis), починаючи із Середньовіччя людям стало відомо, що гібридизація між такими далекими видами переважно неможлива. Тому вважалося, що спадковість і мінливість діє тільки всередині кожного виду, а самі види залишились незмінними від часу їхнього створення[1].
Інше «доменделівське» уявлення про спадковість полягало у тому, що риси успадковуються безпосередньо. Так Гіппократ писав, що репродуктивний матеріал, який він називав «ґонос», від кожної частини тіла батьків незалежно передається відповідній частині тіла дитини і визначає її розвиток. Схожі теорії спадковості були поширені ще до кінця XIX ст. Наприклад, Дарвін 1868 року припускав, що тканини тіла батьків продукують мікроскопічні гранули (gemmules), які передаються дитині і забезпечують формування відповідних структур в ембріона. Уявлення про пряме успадкування переважно поєднувалось із ідеєю про змішування ознак матері і батька. Наприклад, якщо в одного з батьків чорне волосся, а в іншого — біле, діти повинні бути русявими[1].
Суперечливість «класичних» уявлень

«Класичні» уявлення про спадковість, проте, породжували очевидний парадокс: якщо, по-перше, спадковість існує тільки в межах одного виду і немає жодних зовнішніх джерел мінливості, по-друге, риси батьків просто змішуються у потомства, тоді з часом всі особини одного виду мали би стати зовсім однаковими. А це аж ніяк не спостерігається у природі[1].
Перші наукові результати, що ставили під сумнів ідею про пряме успадкування, отримав 1760 року німецький ботанік Йозеф Готліб Кельрейтер. Він схрещував різні сорти тютюну і спостеріг, що перше покоління нащадків відрізнялось від батьківських особин. Проте, якщо схрестити ці рослини між собою, то в наступному поколінні серед усієї різноманітності отриманих рослин можна знайти не тільки дуже схожих до особин першого покоління (своїх батьків), а й до вихідних сортів (своїх «дідусів і бабусь»). Ця робота стала першим кроком до розвитку сучасної генетики. Кельрейтер чітко показав, що, по-перше, ознаки не змішуються, а, по-друге, можуть «маскуватись» в одному поколінні і знову виникати в наступному. Це останнє спостереження очевидно суперечило уявленню про пряме успадкування[1].
Впродовж наступної сотні років робота у напрямку, започаткованому Кельрейтером, продовжувалась. Зокрема, над вдосконаленням культурних рослин працювали деякі англійські джентльмени. Один із них, Т. А. Найт, у 1790-их роках схрестив дві чисті лінії горошку (Pisum sativum) із білими і рожевими квітами. Усі рослини першого покоління мали рожеві квіти, проте у другому знову з'явилась невелика кількість особин із білими пелюстками. Отже, результати Найта підтверджували більш ранні висновки Кельрейтера. Проте, ці роботи мали один вагомий недолік: вони не містили конкретних числових даних. Кельрейтер у своїх пізніх працях писав, що деякі ознаки мають «сильнішу тенденцію» проявлятися, але не пробував описати цю тенденцію математично. Це стало однією із причин того, що «революція» у розумінні спадковості затрималась на століття[1]. Ще одну важливу працю, яка, ймовірно, вплинула на формування ідей Менделя, опублікував у 1863 Чарльз Ноден. У ній французький натураліст обговорює в загальному експерименти із гібридизації рослин, а також повідомляє про розщеплення ознак і домінантність (термін введений Сажере 1826 року) у дурману. Проте Ноден також не описує свої результати математично[2]. Саме базуючись на ідеях попередників, і поєднавши їх із вдалим плануванням експериментів і використанням математичного апарату, Менделю вдалось відкрити основні принципи спадковості.
Досліди Менделя

Грегор Мендель народився 1822 року у Моравії. По закінченню школи і Оломоуцького інституту філософії у віці 21 рік[3] він постригся в монахи Августинського монастиря Святого Томаса у місті Брюнн (зараз Брно, Чехія). У той час в Австро-Угорській імперії політика стосовно релігійних закладів, полягала у тому, що вони повинні були служити не тільки Церкві, а й державі. Через це, монахи мали доступ до великих бібліотек, колекцій гербаріїв, і загалом вели багате інтелектуальне життя[4].
Економічні фактори, такі як розвиток торговельної мережі у Європі, стимулювали розвиток досліджень у галузі сільського господарства. У 1806 році Крістіан Карл Андре заснував Моравське товариство сільського господарства (повна назва: Моравське товариство удосконалення сільського господарства, природничих наук і знань про село). Абат Монастиря Святого Томаса Непп засідав у комітеті цього товариства, а також був президентом Асоціації Помологів і Енологів. Мендель був зацікавлений у вивчені природничих наук і сільського господарства, і абат монастиря його підтримував у цьому. Він посприяв вступу Грегора у Віденський університет, де він вивчав природничі науки і математику, зокрема відвідував курси фізіології рослин Франца Унгера і експериментальної фізики Крістіана Допплера[4]. Не склавши випускних іспитів в університеті Мендель повернувся до монастиря, де провів решту свого життя, і взявся за дослідження гібридизації рослин. Тут у садку монастиря починаючи із 1857 року він проводив досліди, що заклали основи сучасної генетики[5][3]. Свої результати Мендель представив 1865 року на засіданні Брюннського товариства природознавства, а в 1866 році опублікував їх у записках цієї організації[6].
Вибір об'єкта

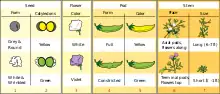
Як модель для своїх досліджень Грегор Мендель вибрав горох посівний (Pisum sativum), той же об'єкт, на якому проводили експерименти Найт та багато інших дослідників. Для цього було кілька причин. По-перше, попередні дані, отримані у працях із цією рослиною, свідчили про те, що можна очікувати розщеплення ознак у потомства. По-друге, у торговців насінням можна було купити різні сорти цієї рослини, що відрізнялись між собою чіткими ознаками, такими як забарвлення віночка або насіння. Мендель почав аналіз і ретельного вивчення 32-ох чистих ліній горошку, і вибрав ті, що відрізнялись за сімома ознаками[7].
По-третє, горошок здатний як до самозапилення, так і до перехресного запилення. Через те, що тичинки і маточка у квітах цієї рослини оточені двома зрослими пелюстками (так званий човен), то пилок із перших переноситься на приймочку останньої, так відбувається самозапилення. Якщо ж дослідник хоче здійснити перехресне запилення, то вирізає тичинки у квітах «материнської» рослини до того, як вони дозріють, і переносить пилок із «батьківської» рослини пензликом або безпосередньо тичинками[8][3].
Окрім того, горошок вигідний із практичних міркувань: він дешевий, невибагливий, не займає багато місця, має відносно короткий цикл розвитку і дає достатньо багато потомства. Тому можна аналізувати одночасну велику кількість рослин і вирощувати кілька поколінь впродовж одного року, що прискорює отримання результатів[8][7].
Особливості підходу
Причиною успіху дослідів Менделя порівняно із його попередниками було вдале планування експериментів. По-перше, він аналізував тільки сім ознак, що мають два чітко відмінні один від одного стани, такі як гладкість/зморшкуватість насіння, зелене/жовте забарвлення сім'ядоль, фіолетові/білі квіти (і, відповідно, сіра/біла шкірка насіння), повні/стягнуті боби, зелене/жовте забарвлення бобів, осьове/термінальне розташування квітів і плодів, високий ріст (180—215 см)/карликовість (20—30 см)[9]. Якби Мендель натомість віддав перевагу дослідженню ознак із неперервним розподілом, таких, як, наприклад, маса насінин, то не зміг би відкрити дискретної природи спадковості. По-друге, він почав свою роботу із того, що перевірив, чи вибрані сорти є справді чистими лініями, тобто чи кожне покоління, отримане від самозапилення, буде зберігати всі батьківські ознаки. На виконання цієї перевірки він затратив два роки[10]. По-третє, він вибрав математичний підхід до опису результатів і обробляв їх статистично.
Постановка типового досліду
Після того, як Мендель пересвідчився, що обрані сорти є чистими лініями, він проводив експерименти із гібридизації. Перші схрещування були моногібридними, тобто такими, у яких враховувалась якась тільки одна ознака, наприклад, колір квітів. Мендель проводив обернені схрещування, тобто, якщо у прямому він переносив пилок із рослин з білими квітами на приймочки рослин із фіолетовими квітами, то в оберненому навпаки. Це робилось для того, щоб пересвідчитись, що походження статевих клітин не впливає на прояв ознаки у нащадків. У випадку тих рис, які аналізував він, обернені схрещування справді давали однакові результати[11].
Після отримання гібридів першого покоління (F1) Мендель дозволяв їм самозапилитись, для того, щоб відбулось розщеплення альтернативних станів ознак у другому поколінні (F2). Після цього він рахував всі рослини із конкретним фенотипом[12]. У деяких дослідах він простежував успадкування ознак до третього покоління.
Аналогічним чином Мендель проводив також і ди- і тригібридні схрещування, у яких батьківські особини відрізнялись за двома чи трьома ознаками відповідно, щоб з'ясувати, як успадкування однієї із них впливає на успадкування іншої. Нарешті, для підтвердження результатів, отриманих у дослідах на горошку, він також проводив схожі експерименти із квасолею (Phaseolus vulgaris і Phaseolus nanus)[13].
Закономірності спадковості, встановлені Менделем
Перший закон
Схрещуючи рослини гороху, що відрізнялись за станами однієї ознаки, Мендель спостеріг, що у фенотипі всіх гібридів першого покоління (F1) проявлявся тільки один із двох станів. Наприклад, всі рослини, отримані внаслідок гібридизації між горохом із білими квітами і горохом із фіолетовими квітами, мали фіолетові квіти. Такі результати підтверджували роботи Найта та інших попередників Менделя і заперечували уявлення про «змішування» ознак батьків у потомства.
Стан ознаки, який проявлявся в F1, Мендель називав домінантним, а той, який не проявлявся — рецесивним. Для всіх семи пар станів ознак, які аналізував Мендель, один виявився домінантним, інший — рецесивним.
| Перший закон Менделя, або закон одноманітності гібридів першого покоління, формулюється так: |
| У першому поколінні від схрещування гомозигот із домінантною та рецесивною ознаками виявляється тільки домінантна ознака[14]. |
Другий закон
Після самозапилення особин F1 Мендель зібрав і висадив насіння із кожної рослини, щоб проаналізувати друге покоління. Цього разу серед рослин з'явились такі, що несли рецесивну ознаку (тобто ту, яка зовсім не виникала в F1). Щоб краще зрозуміти, яким чином відбувається успадкування, Мендель порахував всі особини, у яких проявлялась певна ознака. Наприклад, при схрещуванні гороху із білими і фіолетовими квітами, в F2 було всього 929 рослини, із яких 705 мали фіолетові квіти, і 224 — білі. Результати всіх схрещувань наведені у таблиці[15].
| Фенотип батьків | F1 | F2 | Співвідношення фенотипів у F2 |
|---|---|---|---|
| Кругле × зморшкувате насіння | Всі з круглим насінням | 5474 круглі; 1850 зморшкуваті | 2.96 : 1 |
| Зелене × жовте насіння | Усі із жовтим насінням | 6022 жовті; 2001 зелені | 3.01 : 1 |
| Фіолетові × білі квіти | Усі із фіолетовими квітами | 705 фіолетові; 224 білі | 3.15 : 1 |
| Стягнуті × повні боби | Усі з повними бобами | 882 повні; 299 стягнуті | 2.95 : 1 |
| Зелені × жовті боби | Усі із зеленими бобами | 428 зелені; 152 жовті | 2.82 : 1 |
| Верхівкове × осьове розташування квітів | Усі із осьовими квітами | 651 осьові; 207 верхівкові | 3.14 : 1 |
| Високі × низькі стебла | Усі із високими стеблами | 787 високі; 277 низькі | 2.84 : 1 |
Таким чином в середньому співвідношення фенотипових класів у другому поколінні становило 3:1, тобто у четвертини особин проявлялась рецесивна ознака.
| Другий закон Менделя, або закон розщеплення, говорить: |
| При схрещуванні гібридів першого покоління у нащадків спостерігається розщеплення фенотипових класів у співвідношенні 3:1[14]. |
Після цього Мендель взявся за аналіз особин F2, які проявляли домінантну ознаку, і виявив, що насправді вони не є однорідним класом, а відрізняються одне від одного генетично. У цьому випадку він працював із рослинами, що мали різне забарвлення насіння (жовте — домінантна ознака, зелене — рецесивна). Вибір саме цієї ознаки був зручним через те, що колір кожної горошини визначається її власним генотипом, а не генотипом материнської особини. Тому насінину можна трактувати як окрему особину і оцінювати її фенотип без потреби пророщувати, як це необхідно у випадку багатьох інших ознак. Після схрещування чистих ліній із жовтим і зеленим насінням за описаною вище схемою, Мендель відібрав 519 жовтих горошин F2, проростив їх і дозволив самозапилення. Після цього із кожної рослини окремо зібрав насіння. З'ясувалось, що у 166 особин усе насіння було жовтим, а в решти 353 спостерігалось розщеплення 3:1 між класами із жовтим і зеленим насіння[16].
Провівши аналогічний аналіз особин F2 із зеленими сім'ядолями Мендель пересвідчився, що на відміну від рослин із домінантною ознакою, ці були чистою лінією. Таким чином стало зрозуміло, що за фенотиповим співвідношенням 3:1 криється більш фундаментальне генотипове співвідношення 1:2:1. Це підтвердилось і для інших ознак, які аналізував Мендель. Отже, його модель спадковості пояснювала не розщеплення у другому поколінні 3:1, а саме 1:2:1. Ця модель є прикладом вдалої побудови на основі експериментальних даних наукової гіпотези, що підлягає подальшому дослідному тестуванню[16].
| Третій закон Менделя, або закон незалежного успадкування, говорить: |
| При схрещуванні двох особин, що відрізняються один від одного за двома (і більше) парами альтернативних ознак, гени і відповідні їм ознаки успадковуються незалежно один від одного і комбінуються у всіх можливих поєднаннях (як і при моногібридному схрещуванні). |
Коли схрещувалися гомозиготні рослини, що відрізняються за кількома ознаками, такими як білі і фіолетові квіти і жовте або зелене насіння, успадкування кожної з ознак відповідало першим двом законам і в потомстві вони комбінувалися таким чином, ніби їхнє успадкування відбувалося незалежно одне від одного. Перше покоління після схрещування мало домінантний фенотип за всіма ознаками. У другому поколінні спостерігалося розщеплення фенотипів за формулою 9: 3: 3: 1, тобто 9:16 були з фіолетовими квітами і жовтим насінням, 3:16 з білими квітами і жовтим насінням, 3:16 з фіолетовими квітами і зеленим насінням, 1 : 16 з білими квітами і зеленим насінням.
Менделю трапилися ознаки, гени яких знаходилися в різних парах гомологічних хромосом (нуклеопротеїдних структур в ядрі еукаріотичної клітини, в яких зосереджена велика частина спадкової інформації і які призначені для її зберігання, реалізації і передачі) гороху. При мейозі гомологічні хромосоми різних пар комбінуються в гаметах випадковим чином. Якщо в гамету потрапила батьківська хромосома першої пари, то з однаковою ймовірністю в цю гамету може потрапити як батьківська, так і материнська хромосома другої пари. Тому ознаки, гени яких знаходяться в різних парах гомологічних хромосом, комбінуються незалежно одна від одної. (Згодом з'ясувалося, що з досліджених Менделем семи пар ознак у гороху, у якого диплоїдне число хромосом 2n = 14, гени, що відповідають за одну з пар ознак, перебували в одній і тій же хромосомі. Однак Мендель не виявив порушення закону незалежного успадкування, оскільки зчеплення між цими генами не спостерігалося через велику відстань між ними).
Менделева модель спадковості і її пояснення
Щоб пояснити отримані результати, Мендель запропонував модель спадковості, що складається із кількох припущень, які в результаті лягли в основу класичної генетики.
По-перше, ознаки не передаються безпосередньо від батьків нащадкам, натомість діти успадковують певні дискретні частинки, які несуть інформацію про конкретні ознаки. Пізніше ці часточки спадковості беруть участь у формуванні відповідних рис. Мендель називав їх «факторами», а в сучасній генетиці вони позначаються терміном «ген».
По-друге, кожна особина має по дві копії кожного виду цих часточок спадковості, тобто генів. Вони можуть бути однаковими або відрізнятись. Варіанти гену, які визначають різні стани однієї ознаки, згідно із сучасною термінологією називаються алелями, організми, які несуть два однакові алелі певного гена називають гомозиготними за цим геном, а ті, що несуть різні алелі, — гетерозиготними.
Два алелі одного гену, наявні в однієї особини, не впливають одне на одного, вони не можуть зливатись між собою чи модифікувати одне одного. Через, що залишаються, як висловлюється Мендель, «незабрудненими». Під час утворення статевих клітин (гамет), кожна із них отримує тільки одну із двох наявних у соматичних клітинах «чисту» копію кожного гену. При чому розподіл відбувається порівну: тобто 50 % гамет організму гетерозиготного за певним геном нестимуть один алель, а 50 % — інший. Цей принцип ще називають правилом (законом) чистоти гамет.
| Правило чистоти гамет: |
| Кожна гамета диплоїдного гетерозиготного організму несе лише один алельний ген і не може одночасно містити обидва[17]. |
Запліднення відбувається випадковим чином, тобто гамети комбінуються між собою не в залежності від того, які саме алелі вони несуть. І нарешті, наявність в організму певного алеля ще не гарантує, що він проявиться у фенотипі, у гетерозиготних організмів розвивається тільки один із двох альтернативних станів ознаки, який називають домінантним[18][19].
Перевірка моделі
Менделева модель спадковості чудово пояснювала його експериментальні дані. Проте щоб перевірити її справедливість, необхідно було зробити теоретичні передбачення на основі своєї гіпотези, після чого пересвідчитись, що вони відповідатимуть реальності. Для цього він використав аналізуючі схрещування. Одним із ключових моментів у запропонованій моделі було те, що особини F1 несуть два різні алелі (тобто є гетерозиготами), які рівномірно розділяються між гаметами. Отже, якщо схрестити ці рослини із рослинами гомозиготними за рецесивним алелем, то у наступному поколінні повинне виникнути розщеплення за фенотипом 1:1. Провівши гібридизацію особин F1 із жовтими сім'ядолями, отриманих від схрещування чистих ліній із зеленими і жовтими сім'ядолями, із рослинами з рецесивною ознакою (зеленими сім'ядолями), Мендель отримав розщеплення близьке до 1:1 (58 жовті і 52 зелені)[18][20][21].
Фізичні носії спадковості
Оригінальна стаття Менделя містила багато абстракцій, таких як «фактори спадковості». Крім того, на сучасному йому етапі розвитку біологічної науки не було відомо багатьох процесів і явищ, які дозволили б пояснити механізми домінантності певних алелів, утворення «чистих» гамет тощо. Проте накопичені з того часу знання дозволили підтвердити і доповнити Менделеву модель. Так стало відомо, що «фактори спадковості» або, згідно із сучасною термінологією, гени — це ділянки ДНК, у яких найчастіше закодована інформація, потрібна для синтезу певного поліпептиду (хоча бувають інші типи генів). Різні алелі одного гену відрізняються послідовністю нуклеотидів у них, хоч, зазвичай незначно. Так, на кілька тисяч пар нуклеотидів зміненою переважно є тільки одна або кілька позицій. Для певного гена може існувати велика кількість варіантів, тобто алелів[22].
Домінантність ознак
- Див. також: Домінантність (генетика)
Алелі виникають внаслідок мутацій і їх подальшого закріплення у популяції. Ці мутації часто призводять до того, що продукт гену повністю втрачає свою функціональність. Наприклад, для утворення фіолетового забарвлення пелюсток горошку необхідний пігмент групи антоціанів. Ця хімічна речовина синтезується клітинами у ряді реакцій, що каталізуються ферментами. Останні є білками, інформація про структуру яких закодована у відповідних генах. Рецесивна ознака — біле забарвлення віночка — виникає тоді, коли немає одного із ферментів цього біохімічного шляху. І проявляється вона тільки в тому разі, коли «зламані» обидві копії гену диплоїдного організму (генотип aa), оскільки одного функціонального алеля часто буває цілком достатньо для виконання потрібної роботи. Слід зазначити, що це тільки один із відомих механізмів домінування одних ознак над іншими[22].
Розщеплення ознак
- Див. також: Мейоз
Правило чистоти гамет і закон розщеплення вдалось пояснити після того, як стало зрозуміло, що хромосоми є носіями спадковості у клітинах, і відкриття мейозу. Соматичні клітини багатьох організмів, зокрема горошку і людей, є диплоїдними, тобто містять два набори хромосом, отримані від двох батьківських організмів під час запліднення. Кожна хромосома із пари несе один із двох алелів, при чому розташовані вони в однаковому місці (локусі). Утворенню статевих клітин передує спеціальний тип поділу ядра — мейоз. Під час цього поділу відбувається розділення наборів хромосом. Тобто із утворених внаслідок мейозу гаплоїдних клітин половина міститиме одну хромосому із пари, половина — іншу. Або, іншими словами, для гетерозиготи Aa половина гамет нестиме алель A, половина — a[22].
| ♀\♂ | A | a |
| A | AA | Aa |
| a | Aa | aa |
На основі інформації про те, що різні типи гамет утворюються у гетерозиготи із рівною імовірністю, а запліднення відбувається випадковим чином, можна оцінити імовірність отримання певного фенотипового класу у потомстві. Наприклад, при схрещуванні двох гетерозигот Aa і імовірність появи серед нащадків особин із рецесивною ознакою буде рівна ½ (ймовірність того, що яйцеклітина міститиме алель a) × ½ (ймовірність того, що сперматозоїд міститиме алель a) = 1/4, що відповідає розщепленню 1:3. Такі імовірності можна візуалізувати за допомогою простої діаграми, яка називається решіткою Пеннета. Це двомірна таблиця, у якій у клітинах по горизонталі записують всі можливі типи гамет одного батьківського організму, а по вертикалі — іншого. У клітинках на перетині зазначають можливі комбінації цих гамет, при чому кожна із цих комбінацій має однакову ймовірність виникнення[23].
Незалежне успадкування ознак
- Третій закон або «Закон незалежного успадкування ознак»: кожна пара альтернативних варіантів ознак успадковується незалежно від інших пар і дає розщеплення 3:1 за кожною з пар (як і при моногібридному схрещуванні). При дигібридному схрещуванні (коли спостереження ведеться за двома ознаками) серед гібридів другого покоління спостерігають розщеплення 9:3:3:1. Цей закон справедливий лише для ознак, у яких гени, що їх кодують, належать до різних груп зчеплення, тобто знаходяться в різних хромосомах. Закон може виконуватись і для ознак, гени яких знаходяться в одній хромосомі на значній відстані один від одного (не менше 50 морганід). В іншому випадку гени спадкуватимуться зчеплено.
- Менделю потрапили ознаки, гени яких знаходилися в різних парах гомологічних хромосом (нуклеопротеїдних структур в ядрі еукаріотичної клітини, в яких зосереджена велика частина спадкової інформації і які призначені для її зберігання, реалізації і передачі) гороху. При мейозі гомологічні хромосоми різних пар комбінуються в гаметах випадковим чином. Якщо в гамету потрапила батьківська хромосома першої пари, то з однаковою ймовірністю в цю гамету може потрапити як батьківська, так і материнська хромосома другої пари. Тому ознаки, гени яких знаходяться в різних парах гомологічних хромосом, комбінуються незалежно один від одного. (Згодом з'ясувалося, що з досліджених Менделем семи пар ознак у гороху, у якого диплоїдне число хромосом 2n = 14, гени, що відповідають за одну з пар ознак, перебували в одній і тій же хромосомі. Однак Мендель не виявив порушення закону незалежного успадкування, через те що зчеплення між цими генами не спостерігалося через велику відстань між ними).
Примітки
- Raven et al, 2007, с. 240—241.
- Cook R. A Chronology of Genetics // Year-book of Agriculture. — 1937. — С. 1457—1477.
- Campbell et al, 2008, с. 263.
- Mendel: Man and Mind. Masaryk University Mendel Museum. Архів оригіналу за 16 серпня 2013. Процитовано 13 липня 2013.
- Griffiths et al, 2004, с. 29.
- Sturtevant, 2001, с. 10.
- Raven et al, 2007, с. 242.
- Griffiths et al, 2004, с. 30.
- Tamarin, 2001, с. 19.
- Tamarin, 2001, с. 18.
- Griffiths et al, 2004, с. 31.
- Raven et al, 2007, с. 243.
- Mendel, G., 1866, Versuche über Pflanzen-Hybriden. Verh. Naturforsch. Ver. Brünn 4: 3–47 (in English in 1901, J. R. Hortic. Soc. 26: 1–32)
- Сиволоб та ін., 2008, с. 92.
- Griffiths et al, 2004, с. 32.
- Griffiths et al, 2004, с. 32—33.
- Кучеренко та ін., 2006, с. 50.
- Griffiths et al, 2004, с. 33.
- Raven et al, 2007, с. 247.
- Raven et al, 2007, с. 250.
- Campbell et al, 2008, с. 267.
- Griffiths et al, 2004, с. 34—36.
- Raven et al, 2007, с. 248.
Джерела
- Campbell NA, Reece JB (2008). Biology (вид. 8th). Benjamin Cammings. ISBN 978-0321543257.
- Griffiths A.J.F., Wessler S.R., et al (2004). An Introduction to Genetic Analysis (вид. 8th). W. H. Freeman. ISBN 978-0716749394. Архів оригіналу за 26 лютого 2015. Процитовано 13 серпня 2013.
- Raven P, Johnson G, Singer S, Losos J (2004). Biology (вид. 7th). McGraw-Hill. ISBN 978-0072921649.
- Sturtevant A.H. (2001 (original 1965)). A History of Genetics. Cold Spring Harbor Laboratory Press and Electronic Scholarly Publishing Project. ISBN 0-87969-607-9.
- Tamarin RH (2001). Principles of Genetics (вид. 7th). Mcgraw-Hill. ISBN 0072334193.
- Кучеренко М.Є., Вервес Ю.Г., Балан П.Г., Войціцький П.М. Загальна біологія. — 3-є. — Київ : Генеза, 2006. — ISBN 966-504-199-1.
- Сиволоб А.В., Рушковський С.Р., Кир’яченко С.С. та ін. Генетика: підручник. — Київ : ВПЦ "Київський університет", 2008. — ISBN 975-966-439-108-2.
Посилання
- Менделізм // Українська мала енциклопедія : 16 кн. : у 8 т. / проф. Є. Онацький. — Буенос-Айрес, 1961. — Т. 4, кн. VIII : Літери Ме — На. — С. 953. — 1000 екз.