Крива видового накопичення
Крива́ видово́го накопи́чення (англ. Species discovery curve) — графічне представлення числа видів, знайдених на певній території (або в певному біотопі тощо), як функції кумулятивної сукупності дослідницьких зусиль, спрямованих на їхнє знаходження. Дослідницьке зусилля може вимірюватись як кількість людино-годин спостереження, розставлених пасток, кілометрів буксування тралу судном і т. ін.
Крива видового накопичення є обов'язково зростаючою, і зазвичай це зростання характеризується негативним прискоренням, тобто зростання кривої сповільнюється при додаванні наступної одиниці дослідницького зусилля. В протилежному випадку, коли зростання кривої прискорюється із збільшенням дослідницького зусилля, достовірна оцінка остаточного числа видів є практично неможливою виходячи з міркувань математичної статистики.[1] Побудова такої кривої дозволяє оцінити, скільки нових видів буде виявлено при певному дослідницькому зусиллі; або навпаки, скільки одиниць дослідницького зусилля потрібно для виявлення певного числа видів. Як правило, побудова кривої видового накопичення проводиться методами регресійного аналізу (найчастіше методом найменших квадратів) із використанням нелінійних функцій (див. приклади нижче).
Перше теоретичне обґрунтування кривої видового накопичення та дослідження її властивостей були проведені на матеріалах колекцій малайських метеликів та опубліковані в 1943 році Рональдом Фішером зі співавторами[2].
Незважаючи на широкe застосування методу протягом кількох десятиліть, у низці публікацій він піддається ґрунтовній критиці; зокрема, методами математичного моделювання було показано, що статистично достовірне передбачення загального числа видів у певній фауні за допомогою цього методу можливе лише за умови, якщо наявний перелік вже відомих там видів є близьким до повного.[1]
Приклади описуючих функцій
В наведених нижче функціях S відповідає кількості знайдених видів на кожний момент дослідження, х — дослідницьке зусилля, а, b, c, d — константи, значення яких підбирається при апроксимації, е — основа натурального логарифму.
| (1) |

Логарифмічна модель[2]
| (2) |

Логарифмічна модель для випадку, коли ймовірність знаходження нового виду знижується експоненційно до нуля при збільшенні кількості вже знайдених видів[4]
| (3) |

Рівняння Кленча[5]
| (4) |

Зворотна експоненційна модель для випадку дослідження рідкісних видів[6]
| (5) |

Експоненційна модель для випадку, коли ймовірність знахідки нового виду знижується лінійно до нуля при збільшенні кількості вже знайдених видів[4]
| (6) |

У випадку, коли кількість представників різних видів у досліджуваній сукупності сильно варіює (на порядок величини або більше) рівняння (6) використовують в незначно модифікованій формі (див. приклад в розділі «Приклад застосування методу»):
| (6а) |

Асимптотична регресійна модель[7]
| (7) |

Раціональна функція, використана для емпіричної апроксимації у випадку відсутності теоретичного обґрунтування виду залежності[8]
| (8) |

Модель Чапмана-Річардса[8]
| (9) |

Кумулятивна функція бета-розподілу[9]
| (10) |

Кумулятивна функція розподілу Вейбула[10]
| (11) |

Модель з припущенням про рівну ймовірність знахідки кожного із ще не знайдених видів[11]
| (12) |

У цьому випадку константа репрезентує кількість ще не знайдених видів, — ймовірність повторної знахідки вже знайдених видів при наступному одиничному дослідницькому зусиллі.


В теперішній час для побудови кривих накопичення також активно розробляються та впроваджуються підходи, що ґрунтуються на аналізі та моделюванні стохастичних процесів[12].
При використанні кривої видового накопичення загальна кількість видів на досліджуваній території (в біотопі, таксоні тощо) обчислюється як асимптота функції, використаної для апроксимації даних дослідження; тобто застосовується формула
| (13) |

де N — загальна кількість видів.
Приклад застосування методу
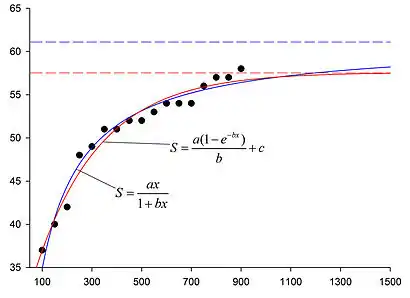
При дослідженні іхтіофауни акваторії острова Зміїний в 2003—2011 роках одним із завдань стала оцінка її видового багатства, тобто загальної кількості видів риб в її складі.[13] Кількість риб різних видів навколо Зміїного відрізняється на 2-3 порядки величини; але різниця між ймовірностями потрапляння в ловчі знаряддя окремо взятого представника кожного з видів є набагато меншою. В такому випадку крива видового накопичення різко зростає на початку дослідження, коли практично одночасно виловлюються представники звичайних видів, а потім, коли до кривої починають додаватись рідкісні види, її зростання сильно уповільнюється (див. рисунок). За таких умов для апроксимації даних польових досліджень використовуються рівняння типу 4 або 6а.
При апроксимації методом найменших квадратів були отримані наступні значення констант апроксимації: для рівняння 4 — а=0.813, b=0.0133; для рівняння 6а — а=0.117, b=0.00386, с=27.263. При таких значеннях констант графіки обох рівнянь показали високу відповідність даним польових досліджень: значення коефіцієнту детермінації R2 дорівнюють 0.9995 та 0.9996 для рівняння 4 та 6а, відповідно.
Після знаходження констант апроксимації їхні значення були підставлені у відповідні рівняння, і згідно з рівнянням (13) обчислена остаточна кількість видів в акваторії (N) при кількості дослідницьких зусиль (х), що прагне до нескінченності; одиницею дослідницького зусилля у цьому випадку вважалася одна постановка ловчого знаряддя. Таким чином були отримані значення N=61.089 для рівняння 4, та N=57.524 для рівняння 6а. Але, при цьому, значення асимптоти для рівняння 6а є меншим, ніж величина останньої точки на емпіричному графіку (див. рисунок), що є неможливим з міркувань загальної логіки.
Ця проблема може бути вирішена шляхом надання більшої статистичної ваги кільком останнім точкам графіку при здійсненні апроксимації кривої;[14] але знаходження конкретної величини коригування статистичної ваги, та обґрунтування кількості точок, статистичну вагу яких треба змінити, є доволі складним завданням. Більш того, коригування даних у такий спосіб означає надання принципово різних властивостей різним точкам експериментального графіку. Це, в свою чергу, суперечить загальному експериментальному підходу, при якому всі дані мають збиратись та оброблятись однаковим методом, і статистичне значення будь-якого одиночного результату (тобто його вплив на остаточний висновок) є однаковим з усіма іншими. Тому, незважаючи навіть на дещо вищу кореляцію графіку рівняння 6а з даними польових досліджень, для опису (апроксимації) цих даних було обране рівняння 4 (рівняння Кленча); і, отже, оцінка загальної кількості видів риб в акваторії о. Зміїний (61 вид) була дана виходячи саме з нього.[13]
Посилання
- Bebber D.P.; Marriott F.H.C., Gaston K.J., Harris S.A., Scotland R.W (2007). Predicting unknown species numbers using discovery curves. Proc. R. Soc. B 274: 1651–8. doi:10.1098/rspb.2007.0464.
- Fisher R.A.; Corbet A.S., Williams, C.B (1943). The relation between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol 42 (1): 42–58.
- Preston F. W (1962). The Canonical Distribution of Commonness and Rarity: Part I. Ecology 43: 185–215. doi:10.2307/1931976.
- Soberόn J.M.; Llorente J.B (2002). The Use of Species Accumulation Functions for the Prediction of Species Richness. Conserv. Biol 7 (3): 480–8. doi:10.1046/j.1523-1739.1993.07030480.x.
- Clench H.K (1979). How to make regional lists of butterflies: some thoughts. J. Lepidopt. Soc 33 (4): 216–31.[недоступне посилання з липня 2019]
- Miller R.I.; Wiegert R.G (1989). Documenting Cmpleteness, Species-Area Relations, and the Species-Abundance Distribution of a Regional Flora. Ecology 70 (1): 16–22. doi:10.2307/1938408.
- Ratkowski, D.A. (1983). Nonlinear regression modeling: a unified practical approach. New York: Marcel Dekker. с. 288. ISBN 0824719077.
- Ratkowski, D.A. (1990). Handbook of nonlinear regression models. New York: Marcel Dekker. с. 241. ISBN 0824781899.
- Mielke P.W.; Johnson E.S (1974). Some generalized beta distributions of the second kind having desirable application features in hydrology and meteorology. Water Resour. Res 10 (2): 223–6. doi:10.1029/WR010i002p00223.
- Brown R.F.; Mayer D.G (1988). Representing cumulative germination. 2. The use of the Weibull function and other empirically derived curves. Ann. Bot 61 (2): 127–38.
- Shen T.-J.; Chao A., Lin C.-F (2003). Predicting the number of new species in further taxonomic sampling. Ecology 84 (8): 798–804. doi:10.1890/0012-9658(2003)084[0798:PTNONS]2.0.CO;2.
- Chao А.; Shen T. J (2004). Nonparametric prediction in species sampling. J. Agr. Biol. Env. Stat 9: 253–69. doi:10.1198/108571104X3262.
- Snigirov S.; Gocharov O., Sylantyev S (2012). The fish community in Zmiinyi Island waters: structure and determinants. Mar. Biodiv. doi:10.1007/s12526-012-0109-4.
- Lamas G.; Robbins R. K., Harvey D.J (1991). A preliminary survey of the butterfly fauna of Pakitza, Parque Nacional del Manu, Peru, with an estimate of its species richness. Publ. Mus. His. Nat. UNMSM 40: 1–19.
|
|
Ця стаття належить до добрих статей української Вікіпедії. |