Цикліофори
Цикліофо́ри (Cycliophora, від грец. cyklion — коло, колесо; phoros — той, що несе) — тип мікроскопічних тварин без порожнини тіл, що живуть як коменсали на ротових органах омарів. Характерною ознакою типу, окрім відсутності целома, є надзвичайно складний життєвий цикл, що поділяється на шість стадій.
? Цикліофори | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
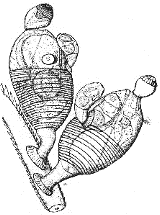 Symbion pandora | ||||||||||||
| Біологічна класифікація | ||||||||||||
| ||||||||||||
Види | ||||||||||||
Посилання | ||||||||||||
| ||||||||||||
Класифікація
Тип містить один рід, в якому на теперішній час описані два види — Symbion pandora та Symbion americanus, та ще один вид відкрито, але наукового опису ще не здійснено. Перелічені види зустрічаються, відповідно, на ротових частинах норвезького омара (Nephrops norvegicus, хазяїн для Symbion pandora), американського омара (Homarus americanus, хазяїн для Symbion americanus), та європейського омара (Homarus gammarus, хазяїн досі не описаного виду). Вперше цикліофори були описані Пітером Фанчем та Рейнхардтом Крістенсеном (Данія) в 1995 році, але ці тварини були відомі зоологам з 1969-го року.
Морфологія
Морфологія цикліофор на різних стадіях життєвого циклу зближує їх з різними групами тварин. Так, ультраструктура кутикули у стадії, що харчується, у великій мірі нагадує кутикулу нематод; формування молодих особин шляхом внутрішнього брунькування зближує цикліофор з мохуватками (Ectoprocta), яким притаманний схожий процес; а будова органу харчування у прикріпленої стадії, і наявність карликових самців, зближує цикліофор з коловертками. Ультраструктурні дослідження епідермісу виявили в його базальному шарі структури, дуже схожі на такі у типу Внутрішньопорошицеві (Entoprocta).
Але складний життєвий цикл цикліофор, котрий включає кілька сидячих та кілька рухливих стадій, не притаманний ані коловерткам, ані мохуваткам та внутрішньопорошицевим; специфічна ж рухлива хордоїдна личинка взагалі є унікальною для первинноротих. Таким чином, еволюційні зв'язки та систематичне положення цикліофор серед тварин досі залишається нез'ясованим.
Будова
Найбільш відомою та вивченою стадією життєвого циклу цикліофор є сидяча особина, здатна до харчування, котра прикріплюється до щетинок на щелепах та ногощелепах омарів. Особини, здатні до харчування, мають близько 350 мікрометрів завдовжки, і складаються із дзвоноподібної ротової воронки, тулуба, та стеблинки із липким прикріплюючим диском. Внутрішня поверхня ротової воронки вкрита війками, а зовнішня війок не несе. Обід ротової воронки складається з приблизно п'ятдесяти війчастих та м'язово-епітеліальних клітин, що чергуються. Війчасті клітини ротової воронки, завдяки своєму биттю, забезпечують захоплення їжі, а м'язові клітини обода, скорочуючись, закривають рот, коли тварина припиняє харчуватись.
Ротова воронка звужується в напрямку до тулуба, і з'єднується з ним вузькою рухливою глоткою. Глотка може бути видовженою або овальною, і створює початок U-подібного харчового тракту, що іде від рота, через глотку проходить до шлунку, потім повертає назад до головного кінця тіла тварини, і закінчується анальним отвором, розташованим поряд з ротовою воронкою. Шлунок цикліофор не має справжнього просвіту, а натомість заповнений масою клітин, які продукують травні ферменти. Стадія, що харчується, не має видільної системи та виражених органів чуття, а нервова система є дуже простою, і складається з маси нервових клітин навколо ротової воронки і пари нервів, що проходять крізь глотку. Ніяких ознак мозку у сидячих особин виявлено не було.
Тулуб цикліофор звужується в напрямку заднього кінця, і переходить у стебельце, котре закінчується диском для прикріплення. І стебельце, і цей диск складаються лише з кутикули, і, таким чином, не мають в своєму складі живих клітин.
Поширення
Розповсюдження Symbion pandora практично повністю збігається з ареалом норвезького омара, який включає в себе Північно-Східну Атлантику з Північним морем та протоками Скагерак та Каттегат. Омари, інфіковані Symbion pandora, були виловлені на Фарерських островах, в Данії, на західному узбережжі Швеції. Також Symbion pandora був знайдений на ротовому апараті норвезьких омарів, виловлених на глибині близько 500 метрів в Середземному морі, але на омарах цього виду, виловлених біля Ісландії та Карибських островів, цього коменсала не знаходять. Symbion americanus був знайдений на відповідному виді омарів (див. вище) біля узбережжя американських штатів Массачусетс та Мен, а третій вид типу — на європейських омарах біля узбережжя Західної Європи.
Спосіб життя
Цикліофори харчуються виключно за рахунок багатого на поживні речовини середовища, що утворюється навколо ротових частин омарів, де особини, здатні до харчування, утворюють щільні колонії. Кількість особин цикліофор залежить від віку омара, і коливається від кількох сотень на молодих ракоподібних до тисяч на старих. Молодий омар линяє частіше, ніж старий, і, оскільки прикріплена популяція комменсалів гине на скинутій кутикулі («панцирі» ракоподібних), кількість цикліофор на старих омарах завжди більша.
Поведінка
Даних про поведінку цикліофор на всіх стадіях життєвого циклу існує дуже мало. Прикріплених особин записували на відео, і ці зйомки показали, що тварини здатні повільно рухати ротовою воронкою, але втягування глотки в тулуб ніколи не спостерігалось. Більшість вільноживучих стадій життєвого циклу, окрім карликових самців та хордоїдних личинок (стадія, що забезпечує розповсюдження), дуже мало здатні до активного плавання.
Живлення
Прикріплена стадія життєвого циклу є єдиною, на якій цикліофори харчуються. Коли омар ловить та поїдає свою здобич, дрібні її частинки та поживні речовини потрапляють у воду навколо його ротового апарату. При битті війок ротової воронки цикліофор створюється ток води, спрямований в глибину воронки. Там поживні частки вловлюються війками безпосередньо, і транспортуються через глотку до шлунку.
Життєвий цикл
Цикліофори мають дуже складний механізм розмноження, з чергуванням статевого розмноження на одній із стадій життєвого циклу, і безстатевого — на двох інших.
Здатні до харчування прикріплені особини не мають ніякої статі, але здатні давати початок іншим, як статевим, так і безстатевим особинам: чоловічим, жіночим, та безстатевим личинкам. Також на цій стадії цикліофори здатні до так званого процесу «самооновлення», в ході якого практично всі живі тканини замінюються новим набором органів. Це самооновлення починається з формування внутрішньої бруньки всередині тіла. Всередині цієї бруньки розвивається повний набір органів, притаманних прикріпленій особині, включаючи травну систему, нервову систему, та ротову воронку. Коли всі органи повністю розвиваються, вони починають повільно зсовують вперед та витісняють старі органи, до тих пір, коли нова ротова воронка проходить крізь глотку та замінює стару. Єдиними частинами старої особини, що залишаються після цього процесу, є оболонка тулубу та прикріплений диск. Сидяча стадія може повторювати цей процес декілька разів, і серед науковців досі немає узгодженої думки, навіщо це робиться. Одне з пояснень може бути таким: позаяк прикріплена стадія не має видільної системи, продукти метаболізму безперервно накопичуються в організмі, і самооновлення дозволяє позбавитись цих продуктів метаболізму до того, як вони почнуть отруювати тварину.
Окрім внутрішніх бруньок в процесі самооновлення, прикріплені особини також можуть давати початок безстатевій нехордоїдній личинці-"пандорі". Ця личинка також розвивається з внутрішньої бруньки, і завжди одночасно з формуванням нового набору органів. Повністю розвинена нехордоїдна личинка виходить із прикріпленої особини через анальний отвір. Вона практично не здатна до плавання, але може повзати по субстрату, використовуючи вкриту війками поверхню на черевній стороні голови. Коли така личинка виходить із материнського організму, вона повільно переповзає на ротові щетинки омара, намагаючись знайти придатне місце для прикріплення, використовуючи чутливі структури, локалізовані на голові. Після знаходження такого місця личинка прикріплюється, личиночні риси негайно дегенерують, і утворюється структура, що нагадує прикріплену цисту.
Коли повзаюча личинка ще знаходиться всередині материнського організму, всередині неї починає розвиватись нова, здатна до харчування особина (і стає видимою в мікроскоп), і ця нова особина містить внутрішню бруньку з яскравими ознаками ротової воронки, що розвивається, яка буде готова до самооновлення відразу після того, як нова прикріплена особина остаточно розвинеться. Коли повзаюча личинка перетворюється на цисту, нова прикріплена особина починає рости у неї всередині, і за короткий час ротова воронка виходить крізь оболонку цисти, після чого нова молода особина починає харчуватись. Формування плазуючих личинок відбувається, переважно, в молодих колоніях, звичайно після линяння омара-хазяїна, коли популяція цикліофор має швидко зростати для відновлення чисельності.
За деяких обставин безстатевий спосіб розмноження може замінюватись на статевий. При статевому розмноженні прикріплені особини можуть продукувати як самиць, так і так званих личинок-прометеїв, які дають початок самцям. Обидва, як і плазуюча личинка-пандора, продукуються при внутрішньому брунькуванні. Личинка-прометей може пересуватись на дуже короткі відстані, і негайно після вивільнення з материнського організму шукає найближчу здатну до харчування особину з жіночою особиною, що розвивається всередині, після чого прикріплюється до цієї материнської особини поблизу її анального отвору. Після прикріплення личинки-прометея в ній починають розвиватись один або два карликові самці. Ці карликові самці є здатними до активного плавання, і добре впізнаються за своїми великими кутикулярними пенісами. Коли жіноча особина виходить з материнського організму через анальний отвір, личинка-прометей вивільняє рухливих самців, котрі швидко знаходять та запліднюють жіночу особину. Жіноча особина виглядає дуже подібно до плазуючої личинки-пандори, але містить всередині один дуже великий ооцит (яйце). Звичайно вона прикріплюється на щупальцях ротового апарату омара, але не всередині колонії здатних до харчування особин. Після прикріплення жіноча особина починає дегенерувати, і формує цисту, як і личинка-пандора. Всередині цисти починає розвиток нова, плаваюча хордоїдна личинка. Ця личинка має велику кількість черевних війок, і є набагато кращим плавцем, ніж будь-яка інша стадія життєвого циклу цикліофор. Таким чином, вона здатна перепливати до інших омарів, або лишатись у воді, доки омар-хазяїн линяє. Коли хордоїдна личинка прикріплюється на ротовому апараті омара, вона перетворюється на цисту, всередині якої починає розвиватись прикріплена, здатна до харчування особина.
Охоронний статус
Ніяких охоронних заходів по відношенню до цикліофор наразі не впроваджується.
Значення
Позаяк цикліофори, будучи дійсними коменсалами, не завдають ніякої шкоди своїм хазяям, вони не мають економічного значення і ніяк не впливають на життя людини.
Посилання
Джерела
- Jessica M. Baker, J. M. and G. Giribet. 2007. A molecular phylogenetic approach to the phylum Cycliophora provides further evidence for cryptic speciation in Symbion americanus. Zoologica Scripta 36(4):353-359.
- Funch, P. 1996. The chordoid larva of Symbion pandora (Cycliophora) is a modified trochophore. Journal of Morphology 230:231-263.
- Funch, P. and R. M. Kristensen. 1995. Cycliophora is a new phylum with affinities to Entoprocta and Ectoprocta. Nature 378:711-714.
- Funch, P. and R. M. Kristensen. 1997. Cycliophora. Pages 409—474 in Microscopic Anatomy of Invertebrates, Vol. 13: *Lophophorates, Entoprocta, and Cycliophora. F. W. Harrison and R. M. Woollacott (eds.) Wiley-Liss, New York.
- Giribet, G., D. L. Distel, M. Polz, W. Sterrer and W. Wheeler. 2000. Triploblastic relationships with emphasis on the acoelomates and the position of Gnathostomulida, Cycliophora, Plathelminthes, and Chaetognatha: a combined approach of 18S rDNA sequences and morphology. Systematic Biology 49:539-562.
- Grzimek's Animal Life Encyclopedia. Vol. 1. Thomson Gale, 2003.
- Kristensen, R.M. 2000. Cycliophora. Pages 251—255 in Encyclopedia of Life Sciences, 5. Macmillan Reference, London.
- Kristensen, R. M. and P. Funch. 2001. Phylum Cycliophora. Pages 199—208 in: Atlas of Marine Invertebrate Larvae. C. M. Young, *M. A. Sewell, and M. E. Rice, eds. Academic Press, London.
- Obst M. and P. Funch. 2003. Dwarf male of Symbion pandora (Cycliophora). Journal of Morphology 255(3):261-278.
- Obst M. and P. Funch. 2006. The microhabitat of Symbion pandora (Cycliophora) on the mouthparts of its host Nephrops norvegicus (Decapoda: Nephropidae). Marine Biology 148(5):945-951.
- Obst M., P. Funch, and R. M. Kristensen. 2006. A new species of Cycliophora from the mouthparts of the American lobster Homarus americanus (Nephropidae, Decapoda). Organisms, Diversity & Evolution 6:83-97.
- Obst, M., P. Funch and G. Giribet. 2005. Hidden diversity and host specificity in cycliophorans: a phylogeographic analysis along the North Atlantic and Mediterranean Sea. Molecular Ecology 14: 4427-4440.
- Winnepenninckx, B., T. Backeljau, and R. M. Kristensen. 1998. Relations of the new phylum Cycliophora. Nature 393:636-638.
