Ядерна пора
Я́дерна по́ра, або нуклеопо́ра — великий білковий комплекс, що проходить крізь нуклеолему (подвійна мембрана, що огортає ядро в клітині еукаріотів), і формує в ній проникний канал. Звичайно на поверхні ядра хребетних знаходиться близько 2000 нуклеопор, але ця кількість може варіювати в широких межах залежно від кількості транскрипцій, що відбулись з клітиною. Білки, що формують ядерну пору, мають назву нуклеопорінів. Близько половини нуклеопорінів містять в своєму складі такі типи складчастої третинної структури, як альфа-соленоїд та бета-пропелер, а в деяких випадках — їх обох. Інша частина нуклеопорінів в своєму складі складчастих структур не мають, тому їм притаманна значна мінливість структури (з огляду на відсутність жорсткої вторинної структури)[1]. Ці невпорядковані протеїни мають назву FG-нуклеопорінів, з огляду на часту повторюванність в їхній первинній структурі пари амінокислот фенілаланін-гліцин (F — однолітерове позначення фенілаланіну, G — гліцину)[2].
Ядерні пори забезпечують транспортування молекул водорозчинних речовин через нуклеолему. Також через нуклеопори можуть траспортуватись такі структури, як РНК (мРНК, компоненти рибосом), білки (ДНК-полімерази, ламіни), вуглеводні, малі молекули, ліпіди. Важливо зауважити, що кожний так званий комплекс ядерної пори (nuclear pore complex, NPC) здатний здійснювати близько 1000 переміщень на секунду. При цьому малі молекули здатні вільно переміщуватись крізь пору завдяки дифузії, а великі мають бути розпізнані за специфічними сигнальними послідовностями, а потім переміщені нуклеопротеїнами всередину ядра або назовні із застосуванням енергопоглинаючих механізмів. Такий механізм переміщення великих молекул відомий під назвою «Ran-ГТФазний цикл» (RAN cycle).
Кожна з восьми білкових субодиниць, що складають оточуюче зовнішнє кільце ядерної пори, має важелеподібну структуру (див. малюнок), що випинається в канал пори. Центральна частина пори частіше за все містить коркоподібне запираюче утворення. Наразі невідомо, чи це утворення є дійсно функціональним запираючим елементом, чи просто масою, що транспортувалася крізь пору, і була зупинена при приготуванні досліджуваних зразків.
Розміри та структура
Весь комплекс нуклеопори має діаметр близько 120 нанометрів, діаметр каналу пори становить близько 50 нанометрів, довжина каналу — приблизно 200 нанометрів. Молекулярна маса нуклеопорного комплексу становить близько 50 МДа і налічує в своєму складі приблизно 30 різних білкових субодиниць[3].
Транспорт через нуклеопору
Об'єкти масою менше 30 кДа здатні проходити крізь нуклеопору завдяки пасивній дифузії. Більші об'єкти теж часом можуть проходити через отвір пори, але в зневажливо малих кількостях[4]. Ефективний транспорт через нуклеопорний комплекс вимагає наявності кількох протеїнових структур[5]. Каріоферіни, котрі можуть виконувати функції як імпортінів (структури, що забезпечують транспорт всередину ядра), так і експортинів (структури, що забезпечують транспорт з ядра назовні) є найбільш вивченими з цих критично важливих структур; всі вони є представниками надродини β-імпортинів, котрим притаманна спільна трьохмірна структура.
На теперішній час існують три моделі, котрі пояснюють механізм транслокації (активного недифузійного переміщення) крізь нуклеопору:
- Градієнтів афінності в напрямку центрального корку
- Броуновського афінного затвору
- Селективних фаз
Імпорт білків
Будь-яка макромолекула, що має в своєму складі ядерну локалізуючу послідовність амінокислот (ЯЛП), здатна зазнавати швидкого транспорту крізь нуклеолему завдяки роботі імпортинів. Відомо кілька ЯЛП; всі вони мають консервативну поліпептидну послідовність з основними амінокислотними залишками, типу PKKKRKV.
Класична схема транспорту ЯЛП-протеїнів починається із зв'язування імпортіну-α з ЯЛП, що забезпечує подальше зв'язування в утвореною структурою імпортіну-β. Комплекс імпортін-α—імпортін-β—протеїн спрямовується до нуклеопори та проходит крізь неї. Після попадання всередину ядра RanГТФ від'єднує від нього імпортін-β. Після цього апоптоз-чутливий клітковий протеїн (cellular apoptosis susceptibility protein, CAS), експортин, що зв'язується з RanГТФ, від'єднує імпортін-α. ЯЛП-протеїн опиняється, таким чином, в нуклеоплазмі вільним. Комплекси імпортін-β—RanГТФ та імпортін-α—CAS—RanГТФ дифундують назад в цитоплазму, де ГТФ гідролізується до ГДФ, що призводить, в свою чергу, до вивільнення імпортіну-α та імпортіну-β, котрі пізніше можуть брати участь в новому транспортному циклі.
Попри те, що транспорт макромолекул крізь ядерну пору відбувається за участі супроводжуючих протеїнів, це переміщення саме по собі не є енергозалежним. Але весь цикл імпорту вимагає гідролізу двох молекул ГТФ; таким чином, він є енергозалежним, і класифікується як різновид активного клітинного транспорту. Цикл імпортування забезпечується наявністю концентраційного нуклео-цитоплазматичного градієнту RanГТФ. Цей градієнт виникає завдяки локалізації виключно в ядрі так званих протеїнів RanGEF, котрі заміщують ГДФ на ГТФ на молекулах Ran. Таким чином, в ядрі, порівняно з цитоплазмою, за нормальних умов наявна підвищена концентрація RanГТФ.
Експорт білків
Декотрі ядерні білки, такі як субодиниці рибосом та РНК-месенджери, синтезуються в ядрі, а потім транспортуються в цитоплазму крізь нуклеопори. Це забезпечується механізмом, що в основних рисах схожий на механізм імпорту.
В класичній схемі експорту протеїни, що містять специфічну ядерну експортну послідовність (ЯЕП) амінокислот, зв'язуються з експортином та RanГТФ, формуючи гетеромерний комплекс. Цей комплекс здатний переміщуватись в цитоплазму шляхом дифузії, після чого ГТФ гідролізується і ЯЕП-протеїн вивільняється. RanГДФ дифундує назад в ядро, де ГДФ замінюється на ГТФ завдяки RanGEF. Цей процес є також енергетично-залежним, позаяк потребує гідролізу ГТФ. Експорт протеїнів за участю екпортинів може бути інгібійований (аж до блокування) антибіотиком лептоміцином.
Експорт РНК
Для кожного з типів РНК існують свої особливі шляхи транспорту з ядра. Цей транспорт також є ЯЕП-залежним, при цьому ЯЕП наявна в протеїнових структурах, з котрими РНК пов'язується при синтезі (за виключенням транспортної РНК, котра не має таких білків-адапторів). Важливо зазначити, що всі типи вірусної РНК та всі типи клітинної РНК, за виключенням матричної, в своєму транспорті залежні від RanГТФ. Деактивована матрична РНК транспортується за участю своїх особливих мРНК-транспортних білкових факторів. Цими факторами є Mex67/Tap (велика субодиниця) та Mtr2/p15 (мала субодиниця). Окрім них, для здійснення процесу транспорту мРНК необхідне зв'язування з великою субодиницею специфічного білкового адаптору.
Додаткові зображення
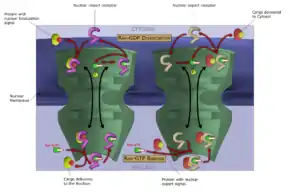 RanГТФ - транспортний цикл
RanГТФ - транспортний цикл
Посилання
- Denning D, Patel S, Uversky V, Fink A, Rexach M (2003). Disorder in the nuclear pore complex: the FG repeat regions of nucleoporins are natively unfolded. Proc Natl Acad Sci U S A 100 (5): 2450–5. PMID 12604785.
- Peters R (2006). Introduction to nucleocytoplasmic transport: molecules and mechanisms. Methods Mol Biol 322: 235–58. PMID 16739728. Архів оригіналу за 28 вересня 2007. Процитовано 16 лютого 2008.
- Alber F, Dokudovskaya S, Veenhoff L, Zhang W, Kipper J, Devos D, Suprapto A, Karni-Schmidt O, Williams R, Chait B, Rout M, Sali A (2007). Determining the architectures of macromolecular assemblies. Nature 450 (7170): 683–94. PMID 18046405.
- Rodriguez M, Dargemont C, Stutz F (2004). Nuclear export of RNA. Biol Cell 96 (8): 639–55. PMID 15519698.
- Reed R, Hurt E (2002). A conserved mRNA export machinery coupled to pre-mRNA splicing. Cell 108 (4): 523–31. PMID 11909523.