Z-ДНК
Z-ДНК — одна з багатьох можливих структур подвійної спіралі ДНК, являє собою лівозакручену подвійну спіраль (на відміну від правозакрученої, як найбільш поширена форма B-ДНК). Z-ДНК є однією з трьох біологічно активних подвійних спіральних структур ДНК, поряд з А-ДНК і В-ДНК, хоча точні її функції до цього моменту не визначені[1].
Історія вивчення
Лівозакручена ДНК вперше була відкрита Робертом Уеллсом і колегами при вивченні полімеру, утвореного повторами дезоксинозин — дезоксицитидін[2]. Вони спостерігали «зворотний» круговий дихроїзм у таких ДНК, з чого зробили вірний висновок, що її ланцюги обвивають один одного в напрямку наліво. Згодом була опублікована кристалічна структура Z-ДНК, де в ході рентгеноструктурного аналізу з'ясувалося, що вона є першим однокрісталлічним фрагментом ДНК (самокомплементарний гексамер ДНК d(CG)3). Було встановлено, що Z-ДНК являє собою лівозакручену подвійну спіраль ДНК з двох антипаралельних ланцюгів, з'єднаних зв'язками між парами азотистих основ. Ці роботи були проведені Ендрю Уонгом (англ. Andrew Wang), Олександром Річем (англ. Alexander Rich) та їх співробітниками в Массачусетському технологічному інституті[3].
У 1970 році було показано, що найбільш поширена B-форма ДНК може переходити в Z-форму. В цьому експерименті було продемонстровано, що круговий дихроізм полімеру (dG-dC) в ультрафіолетових променях в розчині 4М NaCl змінювався на строго протилежний[4]. Те, що при цьому переході В-форма перейшла в Z-форму, було підтверджено результатами рамановскої спектроскопії[5]. Кристалізація сполуки В- і Z-ДНК, проведена в 2005 у[6], дала краще розуміння потенційної ролі, яку Z-ДНК грає у клітині. Скрізь, де є сегменти форм Z-ДНК, повинні бути також В-Z-з'єднання на їх кінцях, що зв'язують Z-форму з B-формою, що зустрічається в усьому іншому геномі.
У 2007 році була описана РНК-версія Z-ДНК як трансформована форма подвійної правозакрученої спіралі A-РНК в лівозакрученій спіралі[7]. Перехід від А-РНК в Z-РНК, тим не менше, був описаний ще у 1984[8].
Структура
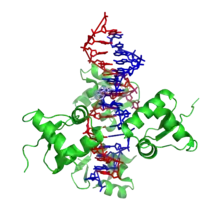
Z-ДНК значно відрізняється від правозакрученних форм. Z-ДНК — лівозакручена і має первинну структуру, повторювану через кожні 2 пари основ. На один поворот спіралі доводиться 12 пар основ. На відміну від А- і В-ДНК, в Z-ДНК велика борозенка слабо помітна, мала борозенка вузька і глибока. Взагалі, структура Z-ДНК енергетично невигідна, хоча деякі умови можуть активізувати її формування, наприклад: пуриново -пиримидинові послідовності (особливо полі (dGC)2) що чергуються, негативна понадспіралізація ДНК, високий вміст солей і деякі катіони (всі при фізіологічній температурі — 37 ° C і pH 7,3-7,4). Z-ДНК може з'єднуватися з B-ДНК у структуру, що приводить до витіснення пар основ (см. Рис.)[9].
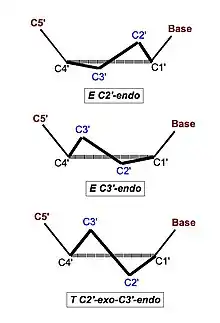
Ще однією особливістю Z-ДНК є чергування конформацій нуклеотидних залишків. Дезоксицитидін знаходиться в стандартній конформації: цукор в С2'-ендоконформаціі (див. малюнок), а азотисті основи — в анти-конформації (тобто основа повернена в сторону, протилежну гідроксильної групи при п'ятому атомі вуглець а; в такому становищі перебувають основи у полінуклеотидному ланцюгу[10]). У дезоксигуанозину цукор знаходиться в С3'-ендоконформаціі, а основа має вкрай нетипову сін- конформацію[11].
Стекінг основ у Z-ДНК володіє новими, властивими лише цій формі, характеристиками. Так, стекінгові взаємодії є тільки між залишками цитозину протилежних ланцюгів, а залишки гуаніну взагалі не взаємодіють один з одним[1].
Фосфати в Z-ДНК не еквівалетні один одному і віддалені на різні відстані від осі спіралі; для гуанінових нуклеотидів ця відстань дорівнює 0,62 нм, а для цитозинових — 0,76 нм. При цьому сусідні цукри «дивляться» в протилежні сторони, і через це лінія, що послідовно з'єднує атоми фосфору в ланцюги, стає зиґзаґоподібною (звідси назва — Z-ДНК)[1] .
Структура Z-ДНК складна для вивчення, тому що вона практично не існує в стабільній формі подвійної спіралі. Навпаки, лівозакручена спіраль Z-ДНК є тимчасовою структурою, що з'являється в результаті біологічної активності і швидко зникає[12].
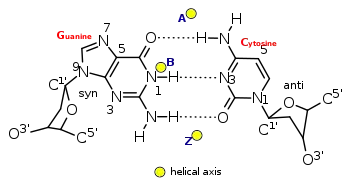
Перехід з В-ДНК в Z-ДНК
Як вже говорилося, В- і Z-форми здатні переходити одна в одну. Це відбувається при зміні іонної сили розчину або концентрації катіонів, що нейтралізують негативний заряд фосфодіефірного каркаса. При цьому для переходу немає необхідності для розходження ланцюгів, він ініціюється розривом водневих зв'язків у кількох пар основ, після чого гуанін фіксується в сін-конформації, водневі зв'язки відновлюються, і основи знову утворюють уотсон-кріковські пари. Область переходу рухається по спіралі у вигляді петлі [1].
Пророкування структури Z-ДНК
На даний момент можливо передбачити правдоподібну послідовність ДНК, що знаходиться у формі Z-ДНК. Алгоритм для передбачення схильності ДНК перебудовуватися з В-форми в Z-форму, ZHunt, був написаний в 1984 році доктором P.Shing Ho з Массачусеткого технологічного інституту[13]. Пізніше цей алгоритм був розвинений Трейсі Кемп і колегами для визначення місць утворення Z-ДНК у всьому геномі[14].
Алгоритм ZHunt доступний за посиланням Z-Hunt online.
Примітки
- конич, Севастьянова, 2012, с. 93.
- Mitsui et al. (1970). Physical and enzymatic studies on poly d (IC) -poly d (IC), an unusual double-helical DNA. Nature 228 (5277): 1166–1169. PMID 4321098.
- Wang AHJ, Quigley GJ, Kolpak FJ, Crawford JL, van Boom JH, Van der Marel G, Rich A (1979). Molecular structure of a left-handed double helical DNA fragment at atomic resolution. Nature 282 (5740): 680–686. Bibcode:1979Natur.282..680W. PMID 514347. doi:10.1038 / 282680a0.
- Pohl FM, Jovin TM (1972). Salt-induced co- operative conformational change of a synthetic DNA: equilibrium and kinetic studies with poly (dG-dC). J. Mol. Biol. 67: 375–396. PMID 5045303. doi:10.1016 / 0022-2836 (72) 90457-3.
- Thamann TJ, Lord RC, Wang AHJ, Rich A (1981). High salt form of poly (dG-dC) • poly (dG-dC) is left handed Z-DNA: raman spectra of crystals and solutions. Nucl. Acids Res. 9: 5443–5457. PMID 7301594. doi:10.1093 / nar / 9.20.5443.
- Ha SC, Lowenhaupt K, Rich A, Kim YG, Kim KK (2005). Crystal structure of a junction between B-DNA and Z-DNA reveals two extruded bases. Nature 437 (7062): 1183–1186. Bibcode:2005Natur.437.1183H. PMID 16237447. doi:10.1038 / nature04088.
- Placido D, Brown BA 2nd, Lowenhaupt K, Rich A, Athanasiadis A (2007). A left-handed RNA double helix bound by the Zalpha domain of the RNA-editing enzyme ADAR1. Structure 15 (4): 395–404. PMC 2082211. PMID 17437712. doi:10.1016 / j.str.2007.03.001.
- Hall K, Cruz P, Tinoco I Jr, Jovin TM, van de Sande JH (October 1984). 'Z-RNA' - a left-handed RNA double helix. Nature 311 (5986): 584–586. Bibcode:1984Natur.311..584H. PMID 6482970. doi:10.1038 / 311584a0.
- de Rosa M, de Sanctis D, Rosario AL, Archer M, Rich A, Athanasiadis A, Carrondo MA (18 травня 2010). Crystal structure of a junction between two Z-DNA helices. Proceedings of the National Academy of Sciences of the United States of America 107 (20): 9088–9092. Bibcode:2010PNAS..107.9088D. PMC 2889044. PMID 20439751. doi:10.1073 / pnas.1003182107.
- конич, Севастьянова, 2012, с. 82.
- конич, Севастьянова, 2012, с. 92.
- Zhang H, Yu H, Ren J, Qu X (2006). Reversible B / Z-DNA transition under the low salt condition and non-B-form polydApolydT selectivity by a cubane-like europium-L-aspartic acid complex. Biophysical Journal 90 (9): 3203–3207. Bibcode:.... 90.3203Z 2006BpJ .... 90.3203Z. PMC 1432110. PMID 16473901. doi:10.1529 / biophysj.105.078402. Архів [http: //www.biophysj.org/cgi/content/full/90/9/3203 оригіналу] за 12 жовтня 2008. Процитовано 21 червня 2019.
- Ho PS, Ellison MJ, Quigley GJ, Rich A (1986). A computer aided thermodynamic approach for predicting the formation of Z-DNA in naturally occurring sequences. The EMBO Journal 5 (10): 2737–2744. PMC 1167176. PMID 3780676.
- Champ PC, Maurice S, Vargason JM, Camp T, Ho PS (2004). [http: //nar.oxfordjournals.org/cgi/pmidlookup? view = long & pmid = 15598822 Distributions of Z-DNA and nuclear factor I in human chromosome 22: a model for coupled transcriptional regulation]. Nucleic Acids Res. 32 (22): 6501–6510. PMC 545456. PMID 15598822. doi:10.1093 / nar / gkh988.
