Геномний імпринтинг
Геномний імпринтинг — це явище, що полягає у відмінності експресії деяких генів в залежності від того, яке вони мають походження — материнське, чи батьківське. Тобто для частини генів активною є тільки копія успадкована від матері (наприклад H19 та CDKN1C), для інших — тільки від батька (наприклад ІФР-2). Геномний імпринтинг — це одна із форм неменделівського успадкування, воно пов'язане із епігенетичними явищами, тобто забезпечується метилюванням ДНК або модифікацією білків гістонів і не супроводжується зміною послідовності нуклеотидів.[1]
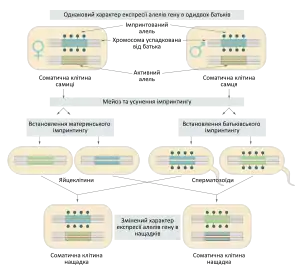
Найбільше прикладів імпринтованих генів було виявлено в організмів, що мають так званий «плацентарний характер» ембріонального розвитку: у плацентраних ссавців та квіткових рослин. Проте окремі випадки знайдені також і в комах (сціарид та плодової мушки), нематод (Caenorhabditis elegans).[2]
Приклади
Синдроми Ангельмана та Прадера-Віллі

%252C_de_Juan_Carre%C3%B1o_de_Miranda..jpg.webp)
Прикладом явища геномного імппринтингу у людини є успадкування синдромів Ангельмана і Прадера-Віллі. Перший проявляється у сповільненні фізичного та розумового розвитку, порушенні сну, приступах, різких рухах, також хворі часто усміхаються, сміються і поводять себе так, наче вони загалом щасливі. Другий синдром полягає у відсутності контролю за споживанням їжі, внаслідок чого хворі часто переїдають і хворіють ожирінням, а також у зниженому тонусі м'язів, сколіозі, підвищеній сонливості, зниженій активності статевих залоз тощо. Обидва синдроми викликаються однією і тією ж мутацією: делецією у ділянці 15q11q13. У 1989 році Ніколлс та колеги з'ясували, що синдром Прадера-Віллі виникає внаслідок делеції у 15-ій хромосомі батьківського походження, в той час як Ангелмана — у хромосомі материнського походження.[3]
Успадкування гену ІФР-2
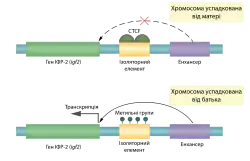
Інший приклад геномного імпринтингу — успадкування гену інсуліноподібного фактору росту 2 (ІФР2), необхідного для пренатального росту (миші, що не експресують цього гену народжуються вдвічі меншими за звичайних). Для фенотипу важливою є тільки батьківська копія цього гену, оскільки тільки вона транскрибується. Таким чином миші, що успадкували дефектний ген від батька будуть карликовими, в той час як дефектний ген успадкований від матері ніяк не впливатиме на розмір потомства.[1]
Імпринтиг батьківського гену ІФР-2 забезпечується метилюванням ДНК, хоча ця модифікація зазвичай викликає пригнічення експресії певного гену, в цьому випадку вона навпаки активує його. Модифікується не сам структурний ген і не його промотор, а так званий ізоляторний елемент поблизу структурного гену. До цієї послідовності в неметильованому стані приєднується білок CTCF, що перешкоджає взаємодії гену ІФР-2 із енхансерною послідовністю. Якщо ізоляторний елемент метильований, то така комунікація стає можливою, через те, що приєднання CTCF не відбувається, відповідно експресія гену ІФР-2 відбувається дуже активно.[1]
Пояснення явища геномного імпринтингу
Диплоїдність вважається еволюційно «вигідною» ознакою, оскільки в організму, що має дві копії певного гену, вони завжди «підстраховують» одна одну, отримавши дефектний ген від одного з батьків особина все ще має шанс на нормальну життєдіяльність завдяки копії отриманій від іншого. Геномний імпринтинг є формою моноалельої експресії генів, за якої описана перевага диплоїдності повністю нівелюється, а отже це явище повинно надавати організмам якусь альтернативну — і сильнішу — селективну перевагу, або мати інше еволюційне пояснення. Біологічна роль геномного імпринтингу досі не з'ясована, на її пояснення було запропоновано кілька гіпотез.
Підвищення здатності до еволюціонування
Одне із пояснень біологічної ролі геномного імпринтингу припускає, що це явище може підвищувати здатність організмів еволюціонувати. Завдяки імпринтингу алель певного гену одне або кілька поколінь може залишатись «в тіні», тобто зовсім не впливати на фенотип організму. За таких умов він не зазнаватиме селекційного тиску, і в ньому зможе накопичитись певна кількість мутацій. Автори гіпотези вважають, що таким чином темпи еволюції можуть збільшитись, оскільки інколи для пристосувальних змін необхідне поєднання двох або більшої кількості мутації, які самі по собі були б летальними.
Ця гіпотеза має ряд недоліків: по-перше, підвищення темпів еволюції вигідне тільки для цілої групи організмів, а не для кожної особини окремо, тому не зрозуміло, яким чином воно могло вперше встановитись у певній групі і що може перешкоджати індивідуумам ревертувати до біалельної експресії генів. По-друге, для алелів із рецесивними мутаціями геномний імпринтинг не тільки не збільшує кількості поколінь, впродовж яких вони не зазнають дії добору, а навпаки — зменшують її. По-третє, ця гіпотеза не дозволяє передбачити які саме гени підлягатимуть імпринтингу і в якої саме статі.[4]
Бомба вповільненої дії яєчників
Вамуза та Менн запропонували на пояснення геномного імпринтингу гіпотезу, яку вони назвали «англ. ovarian time bomb» (OTB) — бомбою вповільненої дії яєчників. Згідно з цим підходом імпринтинг покликаний зменшити імовірність злоякісного переродження клітин зародкової лінії у жінок. Інколи незапліднені ооцити починають спонтанно розвиватись, внаслідок чого виникають яєчникові тератоми, проте, зазвичай такі пухлини є відносно доброякісними, через те, що утворені клітини не можуть диференціюватись в інвазивний трофобласт. Для розвитку трофобласту необхідні гени батька. Автори OTB вважають, що основна роль геномного імпринтингу — «вимкнути» в ооцитах гени, що відповідають за розвиток трофобласту, щоб запобігти утворенню злоякісних тератом. Оскільки у сперматозоїдах ці гени «увімкнуті», після запліднення зможе відбуватись нормальний розвиток.
Хоча ця гіпотеза дає поясення імпринтингу генів, що відповідають за ріст трофобласта, у материнському геномі, і генів, що відповідають за пригнічення трофобластного росту, — у батьківському, проте значення вимкнення не пов'язаних із цими процесами генів, залишається незрозумілим. Автори припускають, що вони могли просто випадково «попасти під гарячу руку» внаслідок неадекватного розпізнавання потрібних генів машинерією імпринтингу. Крім того, ця гіпотеза не може пояснити геномного імпринтингу в організмів із неінвазивною плацентою, наприклад у овець.[4]
Родинна теорія геномного імпринтингу
Найбільше визнання має так звана «родинна теорія» (англ. kinship theory, див. також родинний добір) або теорія конфлікту, яка стверджує, що це явище є наслідком «еволюційної битви статей». Імпринтинг характерний переважно для плацентраних ссавців та квіткових рослин, при чому йому підлягають у великій мірі гени необхідні для ембріонального розвитку. Припускається, що геном батька більше «зацікавлений» у тому, щоб ембріони росли великими (адже з кожна особина намагається поширити свої гени, а великі і здорові діти матимуть більше шансів на виживання), геном матері більше «зацікавлений» у тому, щоб зародки не виростали надто великими, бо це може негативно позначитись на здоров'ї самої самиці. Тому в батьківському геномі ген, що відповідає за ріст увімкнутий, а в материнському — вимкнутий[1].
Основним автором родинної теорії геномного імпринтингу є Девід Гейґ (англ. David Haig), вона базується на концепції включної пристосованості запропонованій Гамільтоном. Згідно з цим підходом пристосувальна цінність гену обраховується як сума ефекту його експресії в організму, що несе цей ген, та ефектів на інші організми помножених на їх спорідненість з особиною носієм. При цьому спорідненість можна поділити на дві складові — спорідненість по материнській лінії (rm) та спорідненість по батьківській лінії (rp), наприклад для зведеного брата по материнській лінії r=1/4, що є середнім арифметичним між rm = 1/2 та rp=0[4].
Якщо розглянути гени, що експресуються у тканинах зародка і впливають на отримання ресурсів від матері, то вони одночасно матимуть позитивний ефект для самої особини (B) і негативний ефект для інших нащадків тієї ж самиці (C). Останні із більшою імовірністю нестимуть алелі, які особина успадкувала від матері, ніж алелі, які вона успадкувала від батька, через те, що самиця може в майбутньому народжувати дітей і від інших самців. Отже в цьому випадку rm > rp, через що алелі успадковані від матері і від батька можуть вступати в конфлікт у питанні, чи брати додаткові ресурси від матері якщо B - rmC < 0 < B - rpC. На це питання можна подивитись з іншої сторони: отримання додаткових ресурсів дає пряму перевагу зародку (B) і коштує його матері (C') та батькові (C). Загалом C' > C і знову ж таки можливий конфлікт між алелями зародка успадкованими від батька та успадкованими від матері, коли B - C' < 0 < B - C[4].
Обидва підходи передбачають, що повинен спостерігатись імпринтинг промоторів росту материнського походження та інгібіторів росту батьківського походження, принаймні за умов, коли самиця може спарюватись із більше ніж одним самцем, більша частина витрат пов'язаних із виведенням потомства лягає саме на неї і нащадки можуть впливати на розподіл материнських ресурсів. Остання умова виконується тільки в організмів із постзиготичною турботою.
Розрахунки в галузі кількісної генетики і теорії ігор показали, що той із алелів (батьківського чи материнського походження), який сприяє більшій кількості продукту певного гену, буде відповідати за всю продукцію цього гену в стані еволюційної рівноваги, інший буде повністю вимкнутий, така особливість отримала назву принципу «гучніший голос торжествує» (англ. loudest-voice-prevails)[4].
Критика родинної теорії
Родинна теорія геномного імпринтигу зазнала критики як стосовно її теоретичної побудови, так і стосовно її застосування для пояснення конкретних феноменів.
Одним із аргументів, що можуть дати підстави сумніватись у логічній цілісності родинної теорії, є те що вона побудована головним чином на основі обрахунків в галузі теорії ігор, в той час як обрахунки популяційної генетики передбачають іншу поведінку алелів генів. Крім того, виникають труднощі із поясненням деяких явищ: наприклад дослідження однобатьківських дисомій (англ. uniparental disomy, UPD), всупереч передбаченням родинної теорії показують, що більшість батьківських UPDs мають результатом зменшений розмір потомства або не мають на нього ніякого ефекту. Також деякі автори вказують на те, що згідно із логікою теорії конфлікту, ефекти імпринтигу мали б зникати після того, як потомство стає незалежним від матері, проте цього не спостерігається[4].
Загалом еволюційне пояснення геномного імпринтингу є полем для активних наукових суперечок, але родинна теорія є найбільш визнаною.
Джерела
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007). Molecular Biology of the Cell (вид. 5th). Garland Science. ISBN 978-0-8153-4105-5.
- Garnier O, Laoueillé-Duprat S, Spillane C (2008). Genomic imprinting in plants. Epigenetics 3 (1): 14–20. PMID 18259119. doi:10.4161/epi.3.1.5554.
- Nicholls RD, Knoll JH, Butler MG, Karam S, Lalande M (1989). Genetic imprinting suggested by maternal heterodisomy in nondeletion Prader-Willi syndrome. Nature 342 (6247): 281–5. PMID 2812027. doi:10.1038/342281a0.
- Wilkins JF, Haig D (2003). What good is genomic imprinting: the function of parent-specific gene expression. Nature reviews, Genetics 4 (5): 359–68. PMID 12728278. doi:10.1038/nrg1062.
Посилання
- Каталог генів, що підлягають імпринтингу в миші на сайті Mousebook.org (англ.)