Клітинний цикл
Клітинний цикл — це проміжок часу від моменту виникнення ядерної клітини до її загибелі або до наступного поділу, тобто серія подій між одним поділом клітини і наступним. Це процес, завдяки якому одна клітина (така як запліднена яйцеклітина) розвивається у сформований організм, і процес, завдяки якому відновлюються волосся, шкіра, клітини крові, а також деякі внутрішні органи. Спеціалізовані форми клітинного поділу відповідають за клітинну диференціацію протягом ембріогенезу і морфогенезу, а також за підтримку стовбурових клітин протягом життя дорослого організму.
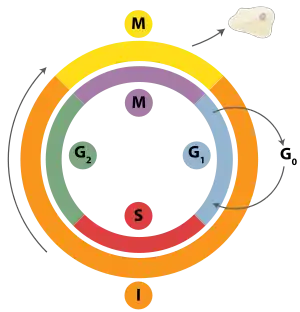
Клітинний цикл складається з чотирьох фаз: фази G1, S-фази, G2-фази (ці три фази також відомі під назвою інтерфаза) і М-фази. М-фаза сама по собі складається з двох тісно пов'язаних процесів: мітозу, коли хромосоми клітини розподіляються між двома дочірніми клітинами, і цитокінезу, коли цитоплазма клітини фізично розділяється. Клітини, що тимчасово припинили ділитися, входять у фазу спокою, так звану G0 фазу. У той самий час клітини, що назавжди припинили поділ через вік або через накопичення пошкоджень у ДНК, називаються старими. Деякі клітини у дорослому організмі, такі як паренхімальні клітини печінки чи нирок, входять у G0-фазу майже назавжди. Вони можуть почати поділ знову тільки за дуже специфічних обставин; інші типи клітин, такі як клітини епітелію, продовжують ділитися протягом усього життя організму.
Вид тканини, до якого належить клітина, зумовлює характер цього циклу в багатоклітинному організмі. В оновлюваних тканинах (епітелій кишок, кістковий мозок, шкіра) 80 % клітин постійно перебувають у мітотичному циклі, а 20 % виходять із циклу, диференціюються та починають виконувати свої специфічні функції.[джерело?] В інших тканинах це співвідношення може бути іншим, наприклад, у печінці й нирках тільки 5-10 % клітин безперервно діляться, а інші виходять з мітотичного циклу й спеціалізуються.
Молекулярні процеси, що відбуваються протягом клітинного циклу, є послідовними. Неможливе «повернення» клітинного циклу у протилежний бік. Існує два ключові класи регуляторних молекул, що спрямовують клітинний цикл: це цикліни і циклін-залежні кінази. Л. Хартвелл (Leland H. Hartwell), Р. Хунт (R. Timothy Hunt) і П. Нерз (Paul M. Nurse) отримали Нобелівську Премію в галузі Медицини і Фізіології 2001 року за відкриття цих центральних молекул у регуляції клітинного циклу.
Фази клітинного циклу
Хоча різні стадії інтерфази не завжди можна розрізнити за морфологією клітини, кожна фаза відрізняється від інших набором біохімічних реакцій, які готують клітину до поділу. Термін «пост-мітотичний» часом уживають як для клітин у стадії спокою, так і для старих клітин. Клітини багатоклітинних еукаріот, коли не діляться, звичайно знаходяться у спокої, у фазі G0, у яку потрапили з фази G1, і можуть залишатися у спокої досить довгий час, навіть невизначено довгий (як здебільшого це відбувається з нейронами). Це досить звична картина для клітин, що остаточно диференційовані. Клітинне ж старіння — це відповідь на пошкодження ДНК або деградацію, що робить нащадків клітини нежиттєздатними. Це просто біохімічна альтернатива саморуйнуванню пошкодженої клітини — апоптозу.
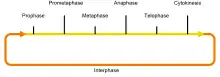
Мітоз
| Складові стадії | Суть складових стадій | Значення в цілому |
|---|---|---|
| Профаза 2n4c | Спіралізація ДНК, утворення подвійних (профазних) хромосом, перехід ядерного матеріалу в хромосоми, руйнування ядерної оболонки | Збільшення кількості клітин і рівномірний розподіл генетичного матеріалу між двома дочірніми клітинами (загалом — процес утворення двох дочірніх клітин, ідентичних материнській клітині — відновлення тканин у процесі їхнього старіння) |
| Метафаза 2n4c | Розміщення подвійних (метафазних) хромосом по екватору, прикріплення хромосом центромерами до ахроматинових ниток, поздовжнє розщеплення хромосом, подвоєння центріолей | |
| Анафаза 2n4c | Руйнування центромерних зв'язок, розходження одинарних (анафазних) хромосом до полюсів, додаткове розходження полярних зон веретена поділу | |
| Телофаза/Цитокінез 2c2c | Деспіралізація хромосом, формування оболонки ядер і ядерця, поділ цитоплазми (цитокінез) та руйнування веретена поділу | |
Мейоз
| Складові стадії | Суть складових стадій | Значення в цілому |
|---|---|---|
| Профаза I | Потовщення, спіралізація, потім — зближення та розміщення поруч гомологічних хромосом (кон'юнгація; внаслідок цього можливий кросинговер) У кінці руйнується ядерна оболонка, утворюється веретено поділу та відбувається попарне переміщення хромосом до екватора клітини | Забезпечення поділу статевих клітин (утворення чотирьох гамет з гаплоїдним набором хромосом з вихідної материнської клітини з диплоїдним набором хромосом) |
| Метафаза I | Попарне становлення над і під площиною екватора гомологічних хромосом | |
| Анафаза I | Розходження гомологічних хромосом до полюсів клітини (кількість генетичного матеріалу зменшується вдвічі) | |
| Телофаза I | Утворюються дві клітини або два ядра з гаплоїдним набором хромосом, початок другого поділу | |
| Профаза II | Схожий до профази мітозу та найчастіше не відбувається | |
| Метафаза II | Перехід гомологічних хромосом, які складаються з двох хроматид, до екватора клітини | |
| Анафаза II | Розходження хроматид до протилежних полюсів | |
| Телофаза II | Схожа до телофази мітозу: утворення чотирьох клітин з гаплоїдним набором хромосом | |
Амітоз
| Складові стадії | Суть складових стадій | Значення в цілому |
|---|---|---|
| Одна стадія зумовлює поділ ядра спеціалізованих тканин, приречених на загибель | Поділ ядра, та, іноді, цитоплазми, без утворення хромосом та веретена поділу; приблизне розподілення хромосомного набору | Збільшення кількості клітин, утворення багатоядерних клітин та нерівномірний розподіл матеріалу кожної хромосоми між дочірніми клітинами (спеціалізація клітин) |
Інтерфаза
| Складові стадії | Суть складових стадій | Значення в цілому |
|---|---|---|
| Пресинтетичний період G1 фаза | Інтенсивний синтез білка та ріст клітини | Зумовлює умови для періоду між поділами клітин (загалом — період між поділами клітин) |
| Синтетичний період S-фаза | Редуплікація ДНК, синтез білків хромосом, потовщення хромосом | |
| Постсинтетичний період G2 фаза | Інтенсивна підготовка до мітозу | |
| Кінцева диференціація G0 фаза | Клітини виходять з клітинного циклу і постійно залишаються в інтерфазі, без поділу. | Клітини виконують свої функції. Прикладами таких клітин є нейрони, кардіоміоцити. |
Цикліни і циклін-залежні кінази
Цикліни і циклін-залежні кінази, Cyclins and cyclin-dependent kinases (CDKs), це два класи молекул, необхідних для регуляції клітинного циклу. Цикліни формують регуляторну субчастину, а кінази — каталітичну субчастину активованого гетеродимеру. Цикліни не мають ферментативної активності самі по собі, тоді як циклін-залежні кінази не можуть бути активними без взаємодії з циклінами. У стані гетеродимеру з циклінами циклін-залежні кінази каталізують звичайну біохімічну реакцію — фосфорилювання. Зазвичай, залишок фосфорної кислоти відщеплюється від молекули АТФ (гамма-фосфат у цьому випадку) і переноситься на білок-мішень, що призводить до активації чи дезактивації цієї мішені. Завдяки цьому направляється перехід клітини до наступної фази клітинного циклу.
Більшість із тих генів, що кодують цикліни і циклін-залежні кінази, є консервативними у всіх еукаріотів. Проте, як правило, у складнішого організму і система регуляції клітинного циклу є складнішою. Більшість генів, пов'язаних із клітинним циклом, була вивчена на дріжджах, особливо на Saccharomyces cerevisiae. Номенклатура генів у цих організмів складалася таким чином: до cdc (тобто цикл клітинного поділу, «cell division cycle»), додавали якийсь ідентифікаційний номер. Наприклад, cdc25.
Таким чином, одна і та ж молекула може бути представлена у вигляді гомологів у досить різних організмах. При цьому, деякі організми можуть уживати одну й ту ж молекулу для різних цілей, можуть комбінувати різні функції в одній молекулі.
Після одержання про-мітотичного сигналу ззовні (екстраклітинний сигнал) G1 комплекс циклін — циклін-залежна кіназа активується і готує клітину до S-фази, забезпечуючи експресію транскрипційних факторів, які, у свою чергу, забезпечують експресію циклінів S-фази і білків, необхідних для реплікації ДНК. G1 комплекс циклін — циклін-залежна кіназа також забезпечує деградацію білків — інгібіторів переходу до S-фази шляхом убіквітинування. Білки, після приєднання до них молекул убіквітину, стають мішенню для протеасоми — комплексу білків, що забезпечує протеолітичну деградацію, або протеоліз. Активований комплекс циклін — циклін-залежна кіназа також фосфорилює білки, з яких складається пре-реплікаційний комплекс G1-фази на оріджинах реплікації ДНК, тобто на тих місцях ДНК, з яких, власне, і починається процес реплікації. Фосфорилювання тут має дві мети: перше — це перетворити неактивний пре-реплікаційний комплекс на активний, і друге — це запобігти формуванню нових пре-реплікаційних комплексів. Завдяки цьому кожна частина геному клітини буде реплікована (подвоєна) один і тільки один раз. Останнє є дуже важливим для наступних поколінь клітин. Звичайно, дочірні клітини, у яких не буде частини геному, скоріше за все, помруть. Але й мутації, викликані більш ніж одним поділом якоїсь частини геному, скоріше, будуть шкідливими.
Мітотичні комплекси циклін — циклін-залежна кіназа, які були синтезовані протягом S-фази і G2-фази, але знаходилися у неактивованому стані до власне стадії мітозу, забезпечують ініціацію мітозу, стимулюючи білки, які відповідають за конденсацію хромосом і формування мітотичного веретена. Важливим білковим комплексом, що активується у цей час, є убіквітин-лігаза, відома як комплекс, що забезпечує анафазу (анафаза — одна зі стадій мітозу). Ця убіквітин-лігаза забезпечує, у свою чергу, деградацію структурних білків, які пов'язані з хромосомним кінетохором. Цей убіквітин-лігазний комплекс також забезпечує деградацію мітотичних циклінів, і, таким чином, перехід до телофази мітозу і цитокінезу.
Джерела
- Bruce Alberts, Alexander Johnson, Julian Lewis, David Morgan, Martin Raff Keith Roberts, and Peter Walter (2014). Molecular biology of the cell (англ.) (вид. 6). Garland Science. ISBN 9780815344322. OCLC 902708011.
- А. В. Сиволоб, С.Р. Рушковський, С.С. Кир'яченко та ін. (2008). Генетика. К: Видавничо-поліграфічний центр "Київський університет". с. 47-52.