Синаптотагмін
Синаптотагмін (англ. synaptotagmin) - трансмембранний кальцій-зв'язуючий білок, який відіграє важливу роль у внутрішньоклітинному мембранному транспорті.
Відкриття
Використовуючи методи біохімії та молекулярної біології, лауреат Нобелівської премії з фізіології або медицини Томас Зюдгоф (Стенфордський університет, США) ідентифікував білки, що відіграють важливу роль у роботі синаптичних везикул. Він показав, що один з білків везикулярної мембрани — синаптотагмін — має два окремих домени: один для зв'язування з кальцієм, інший — з фосфоліпідним шаром мембрани. Це вказало на його потенційну значимість для вивільнення нейромедіатора, тому що підвищення внутрішньоклітинної концентрації кальцію супроводжує активацію нейрона. За допомогою методів створення трансгенних тварин Зюдгоф з'ясував функціональну роль синаптотагміна і декількох інших везикулярних білків, необхідних для швидкого викиду нейромедіатора у відповідь на кальцієвий сигнал.
Структура та функції
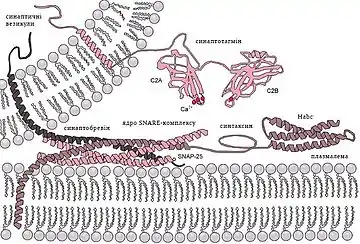
В основі секреції медіатора в хімічних синапсах лежить процес екзо- та ендоцитозу синаптичних везикул в активних зонах. Екзоцитоз синаптичних везикул здійснюється за участю ряду білків, в тому числі синтаксинів, які в даному процесі відіграють ключову роль. У цьому процесі синтаксини взаємодіють з групою білків синаптотагмінів мембрани синаптичних везикул. Синтаксини і синаптотагміни є головними Са2+-сенсорами екзо- і ендоцитозу, просторово сполученими з Са2+-каналами.
Синаптотагмін у людини та інших ссавців представлений 17 ізоформами білка, що кодуються окремими генами: SYT1, SYT2, SYT3, SYT4, SYT5, SYT6, SYT7, SYT8, SYT9, SYT10, SYT11, SYT12, SYT13, SYT14, SYT15, SYT17.
Молекула синаптотагміна складається з N-кінцевого трансмембранного фрагмента, зв'язуючого елементу, та двох C2 доменів (C2A і C2B), що зв'язують кальцій. Синаптотагмін є кальцієвим сенсором, який бере участь в останніх стадіях викиду нейромедіатора в синаптичну щілину. При цьому він зв'язується з нейрексином-1α[2] і SNAP25[3], здійснюючи утримання секреторної везикули біля пресинаптичної мембрани, і бере участь у викиді нейромедіатора за рахунок регуляції SNARE-комплексу при підйомі концентрації кальцію.
Зв'язуючись з кальцієм в певному інтервалі концентрацій, синаптотагмін змінює ступінь своєї взаємодії з білками синаптичних везикул і плазматичної мембрани. У мутантних тварин, у яких відсутній ген, що кодує синаптотагмін, стимуляція аксона не надає ефекту або має незначний вплив на вивільнення медіаторів, хоча спонтанне вивільнення медіаторів підвищено. Таким чином, синаптотагмін діє як регулятор, що запобігає зливанню мембран за відсутності кальцію і підсилює злиття мембран і екзоцитоз при надходженні кальцію всередину клітини.
С-кінцевий С2 домен
Показано, що С-кінцевий С2 домен синаптотагміну складається з 130-140 амінокислотних залишків. Також на прикладі синаптотагміну 1 було доведено, що С2 домен служить для зв'язування кальцію. Дослідження синаптотагміну-1 за допомогою ядерного магнітного резонансу (ЯМР) показали, що кальцій зв'язується виключно з верхньою петлею, і цей процес координується залишками аспартату: три іони кальцію зв'язуються з C2A доменом через D172, D178, D230, D232,D235 і D238 і два іони кальцію зв'язується з C2B доменом через D303, D309, D363, D365 і D371. Не всі ізоформи синаптотагміну у домені С2 зв'язують іони кальцію, а лише Syt1, Syt2, Syt3, Syt5, Syt6, Syt7, Syt9, Syt10. Ці вісім ізоформ синаптотагміну задовольняють потреби у зв'язуванні кальцію для регулювання екзоцитозу. Інші сім ізоформ не зв'язуються з кальцієм у зв'язку з нестачею кислотних залишків, що координують зв'язування, або з специфічною їх просторовою орієнтацією.
Домен С2А регулює злиття синаптичних везикул під час екзоцитозу[4][5] Також кінетика Ca2+-залежного зв'язування фосфоліпідів доменом C2A in vitro збігається з дуже швидкою механікою вивільнення нейромедіаторів (впродовж 0,2 с).[6]. Домен C2B зв'язується з фосфатидил-інозитол-3,4,5-трифосфатом при відсутності іонів кальцію і фосфатидил-інозитол-бісфосфатом в їх присутності, припускають, що ліпідні взаємодії відбувається під час деполяризації клітинної мембрани.
Нещодавно було показано, що синаптотагмін-1 може витісняти комплексин з SNARE-комплексу в присутності кальцію. Вважається, що це один з останніх кроків у процесі екзоцитозу[7].
Посилання
- Georgiev, Danko D.; James F. Glazebrook (2007). Subneuronal processing of information by solitary waves and stochastic processes. У Lyshevski, Sergey Edward. Nano and Molecular Electronics Handbook. Nano and Microengineering Series. CRC Press. с. 17–1–17–41. ISBN 978-0-8493-8528-5. Архів оригіналу за 16 січня 2016. Процитовано 10 квітня 2013.
- Fukuda M, Moreira JE, Liu V, Sugimori M, Mikoshiba K, Llinas RR (2000). Role of the conserved WHXL motif in the C terminus of synaptotagmin in synaptic vesicle docking. Proc Natl Acad Sci USA 97 (26): 14715–14719. PMC 18984. PMID 11114192. doi:10.1073/pnas.260491197.
- Schiavo G, Stenbeck G, Rothman JE, Söllner TH (1997). Binding of the synaptic vesicle v-SNARE, synaptotagmin, to the plasma membrane t-SNARE, SNAP-25, can explain docked vesicles at neurotoxin-treated synapses. Proc Natl Acad Sci USA 94 (3): 997–1001. PMC 19628. PMID 9023371. doi:10.1073/pnas.94.3.997.
- . Відповідно до цього, кінетика кальцій-залежних фосфолипідів забезпечує активність домену C2A in vitro це складає дуже швидкий характер вивільнення нейромедіаторів (у межах 200 мкс). Zimmerberg J, Akimov SA, Frolov V (2006). Synaptotagmin: fusogenic role for calcium sensor?. Nature Structural & Molecular Biology 13 (4): 301–303. PMID 16715046. doi:10.1038/nsmb0406-301.
- Fernández-Chacón R, Königstorfer A, Gerber SH, García J, Matos MF, Stevens CF, et al. (2001). Synaptotagmin I functions as a calcium regulator of release probability. Nature 410 (6824): 41–49. PMID 11242035. doi:10.1038/35065004.
- Chapman ER (2002). Synaptotagmin: A Ca2+ sensor that triggers exocytosis?. Nature Reviews Molecular Cell Biology 3 (7): 498–508. PMID 12094216. doi:10.1038/nrm855.
- Tang J, Maximov A, Shin OH, Dai H, Rizo J, Südhof TC (2006). A complexin/synaptotagmin 1 switch controls fast synaptic vesicle exocytosis. Cell 126 (6): 1175–1187. PMID 16990140. doi:10.1016/j.cell.2006.08.030.
Джерела
- Lang T Jahn R (2008). «Core proteins of the secretory machinery». Handb Exp Pharmacol (184): 107–27.
- Ute Becherer & Jens Rettig: Vesicle pools, docking, priming, and release Cell Tissue Res (2006) 326:393–407.
