Фотосинтез
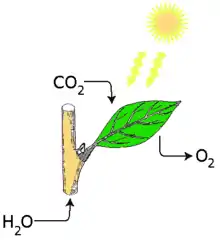
Фотоси́нтез (від грец. φωτο- — світло та грец. σύνθεσις — синтез, сукупність) — процес синтезу органічних сполук з вуглекислого газу та води з використанням енергії світла й за участю фотосинтетичних пігментів (хлорофіл у рослин, хлорофіл, бактеріохлорофіл і бактеріородопсин у бактерій), часто з виділенням кисню як побічного продукту. Це надзвичайно складний процес, що передбачає довгу послідовність узгоджених біохімічних реакцій. Він відбувається у вищих рослинах, водоростях, багатьох бактеріях, деяких археях і найпростіших — організмах, відомих разом як фототрофи. Сам процес відіграє важливу роль для кругообігу вуглецю у природі.
.jpg.webp) |
| Паливо |
| Фізичні основи |
|
Сонце · Сонячна радіація |
| Викопне паливо |
|
Вугілля · Горючі сланці · Гідрат метану · Нафта · Природний газ · Торф |
| Природне невикопне паливо |
|
Водорості · Деревина · Рослинні і тваринні жири та олії · Трава |
| Штучне паливо |
|
Біопаливо · Генераторні гази · Кокс · Моторні палива |
| Концепції |
|
Енергетична біосировина |
Фотосинтез — єдиний процес у біосфері, який призводить до засвоєння енергії Сонця і забезпечує існування як рослин, так і всіх гетеротрофних організмів.[1]
Рослина поглинає не всю сонячну енергію, а лише її фотосинтетичну активну радіацію, під впливом якої проходить фотосинтез, що впливає на майбутній врожай культур.[2]
Узагальнене рівняння фотосинтезу (брутто-формула) має вигляд:
- 6СО2 + 6Н2О = С6Н12О6 + 6О2
Типи фотосинтезу
Розрізняють оксиґенний і аноксиґенний типи фотосинтезу. Оксиґенний найбільш поширений, його здійснюють рослини, ціанобактерії і прохлорофіти. Аноксиґенний фотосинтез проходить у пурпурних, деяких зелених бактеріях та геліобактеріях.
Вирізняють три етапи фотосинтезу: фотофізичний, фотохімічний та хімічний. На першому етапі відбувається поглинання фотонів світла пігментами, їх перехід в збуджений стан і передавання енергії до інших молекул фотосистеми. На другому етапі відбувається розділення зарядів в реакційному центрі, перенесення електронів фотосинтетичним електроно-транспортним ланцюгом, що закінчується синтезом АТФ і НАДФН. Перші два етапи разом, називають світлозалежною стадією фотосинтезу. Третій етап відбувається вже без обов'язкової участі світла і охоплює біохімічні реакції синтезу органічних речовин з використанням енергії, накопиченої на світлозалежній стадії. Найчастіше як такі реакції, розглядається цикл Кальвіна і глюконеогенез, утворення цукрів і крохмалю з вуглекислого газу повітря.
Просторова локалізація
Фотосинтез рослин здійснюється в хлоропластах, відособлених двомембранних органелах клітини. Хлоропласти можуть бути в клітинах плодів, стебел, проте основним органом фотосинтезу, анатомічно пристосованим до його здійснення, є листя. У листку найбагатша хлоропластами тканина — палісадна, або фотосинтезуюча/стовпчаста/хлорофілоносна, паренхіма. У деяких сукулентів з виродженим листям (наприклад, кактусів) основна фотосинтетична активність пов'язана зі стеблом.
Світло для фотосинтезу захоплюється повніше завдяки плоскій формі листка, що забезпечує велике відношення поверхні до об'єму. Вода доставляється з кореня розвиненою мережею судин (прожилків листка). Вуглекислий газ надходить частково за допомогою дифузії через кутикулу і епідерміс, проте велика його частина дифундує в листя через продихи і по листку через міжклітинний простір. Рослини, що здійснюють С4 і CAM-фотосинтез, сформували особливі механізми для активної асиміляції вуглекислого газу.
Внутрішній простір хлоропласта заповнений безбарвною речовиною (стромою) і пронизаний мембранами (ламелами), які, з'єднуючись один з одним, утворюють тилакоїди, які у свою чергу групуються в стопки, так звані грани. Внутрішньотилакоїдний простір відокремлений і не сполучається з рештою строми, передбачається також, що внутрішній простір всіх тилакоїдів сполучений між собою. Світлові стадії фотосинтезу пов'язані з мембранами, автотрофна фіксація вуглекислого газу відбувається в стромі.
У хлоропластах є власні ДНК, РНК, рибосоми (типу 70S), де йде синтез білка (хоча цей процес і контролюється з ядра). Вони не синтезуються знову щоразу, а утворюються шляхом поділу попередніх. Все це дозволило вважати їх попередниками вільних ціанобактерій, що увійшли до складу еукаріотичної клітини в ході симбіогенезу.
Ціанобактерії та інші фотосинтезуючі бактерії та археї, таким чином, самі виконують функції хлоропластів рослин, і фотосинтетичний апарат їх клітин не винесений в особливу органелу. Їхні тилакоїди, проте, не утворюють стопок, а утворюють різні складчасті структури (в однієї ціанобактерії Gloeobacter violaceus тилакоїди відсутні зовсім, а весь фотосинтетичний апарат знаходиться на цитоплазматичній мембрані, що не створює заглибин). У них і в рослин також є відмінності у світлозбиральному комплексі (див. нижче) і в складі пігментів.
Світлова (світлозалежна) стадія
В ході світлової стадії фотосинтезу утворюються високоенергетичні продукти: аденозинтрифосфат, який служить в клітині джерелом енергії, і НАДФН, що використовується як відновник. Як побічний продукт виділяється кисень.
Фотохімічна суть процесу
Хлорофіл має два рівні збудження (з цим пов'язана наявність двох максимумів на його спектрі поглинання): перший пов'язаний з переходом на вищий енергетичний рівень електрона системи зв'язаних подвійних зв'язків, другий, — зі збудженням неспарених електронів азоту і кисню порфіринового ядра. При незмінному спіні електрона формуються синглетні перший і другий збуджений стан, при зміненому — триплетні перший і другий.
Другий збуджений стан найбільш високоенергетичний, нестабільний, і хлорофіл за 10−12 с переходить з нього на перший, з втратою 100 кДж/моль енергії лише у вигляді тепла. З першого синглетного і триплетного станів молекула може переходити в основний з виділенням енергії у вигляді світла (флуоресценція) або тепла, з перенесенням енергії на іншу молекулу, або, оскільки електрон на високому енергетичному рівні слабко зв'язаний з ядром, з переносом електрона на іншу сполуку.
Друга можливість здійснюється у світлозбиральних комплексах, перша — в реакційних центрах, де переведений до збудженого стану при поглинанні кванта світла хлорофіл, стає донором електрона (відновлювачем) і передає його на первинний акцептор. Щоби запобігти поверненню електрона на позитивно заряджений хлорофіл, первинний акцептор передає його вторинному. Крім того, час життя отриманих сполук вищий, ніж у збудженої молекули хлорофілу. Відбувається усталення енергії і розділення зарядів. Для подальшої стабілізації вторинний донор електронів окислює позитивно заряджений хлорофіл, первинним же донором у разі оксигенного фотосинтезу, є вода.
Проблемою, з якою стикаються при цьому організми з оксигенним типом фотосинтезу, є відмінність окислювально-відновних потенціалів води (E0=+0,82 В) і НАДФ+ (E0=-0,32 В). Хлорофіл при цьому повинен мати в основному стані потенціал, більший за +0,82 В, щоб окиснювати воду, але при цьому мати у збудженому стані потенціал менший ніж −0,32 В, щоби відновлювати НАДФ+. Одна молекула хлорофілу може відповідати обом вимогам завдяки наявності ізольованих систем спряжених π-зв'язків: акцептором електронів у молекулах хлорофілів виступає вільна високоенергетична d-орбіталь атома магнію, спряжена з ароматичною системою порфінового ядра, донором виступає карбоксиметильна група, приєднана до атома вуглецю в положенні 13² (нумерація на сторінці хлорофіл німецькою), яка має зв'язок з карбонільною групою, приєднаною до атома вуглецю в положенні131. Перенесення електрона від карбонільної до карбоксиметильної групи відбувається крізь водневий атом координаційно зв'язаної молекули води (або атом кетальної форми), після чого водневий зв'язок послаблюється або розривається через набуття карбоксиметильною групою порівняно добре локалізованого позитивного заряду. Таким чином, запобігання втраті поглиненої енергії через флуоресценцію вдається досягти внутрішньомолекулярною дисоціацією.
Світлозбиральні комплекси
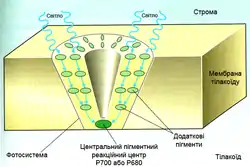
Хлорофіл виконує два завдання: поглинання і передавання енергії. Більше 90 % всього хлорофілу хлоропластів входить до складу світлозбиральних комплексів (СЗК), що виконують роль антени, яка передає енергію до реакційного центру фотосистем I або II. Крім хлорофілу, в СЗК є каротиноїди, а у деяких водоростей і ціанобактерій — фікобіліни, роль яких полягає в поглинанні світла тих довжин хвиль, котрі хлорофіл поглинає порівняно слабо.
Передавання енергії йде резонансним шляхом (механізм Лерстера) і посідає для однієї пари молекул 10−10–10−12 с, відстань, на яку здійснюється перенесення, становить близько 1 нм. Передавання супроводжується деякими втратами енергії (10 % від хлорофілу а до хлорофілу b, 60 % від каротиноїдів до хлорофілу), через що можлива тільки від пігменту з максимумом поглинання за меншої довжини хвилі до пігменту з більшою довжиною хвилі в максимумі поглинання. Саме у такому порядку взаємно локалізуються пігменти СЗК, причому найбільш довгохвильові хлорофіли знаходяться в реакційних центрах. Зворотний перехід енергії неможливий.
СЗК рослин розташований в мембранах тилакоїдів, у ціанобактерій основна його частина винесена за межі мембран у прикріплені до них фікобілісоми — паличкоподібні поліпептидно-пігментні комплекси, в яких знаходяться різні фікобіліни: на периферії фікоеритрини (з максимумом поглинання при 495—565 нм), за ними фікоціаніни (550—615 нм) і алофікоціаніни (610—670 нм), що послідовно передають енергію на хлорофіл а (680—700 нм) реакційного центру.
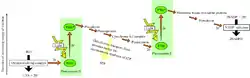
Фотосистема II
Фотосистема — сукупність СЗК, фотохімічного реакційного центру і переносників електрона. Світлозбиральний комплекс II містить 200 молекул хлорофілу а, 100 молекул хлорофілу b, 50 молекул каротиноїдів і 2 молекули феофітину. Реакційний центр фотосистеми II є пігмент-білковим комплексом, розташованим в тилакоїдних мембранах і оточеним СЗК. У ньому знаходиться димер хлорофілу а з максимумом поглинання при 680 нм (П680). На нього кінець-кінцем передається енергія кванта світла з СЗК, внаслідок чого один з електронів переходить у вищий енергетичний стан, зв'язок його з ядром ослаблюється, і збуджена молекула П680 стає сильним відновником (E0=-0,7 В).
П680 відновлює феофітин, надалі електрон переноситься на хінони, що входять до складу ФС II, і далі на пластохінони, транспортується у відновленій формі до b6f-комплексу. Одна молекула пластохінона переносить 2 електрони і 2 протони, які беруться зі строми. Заповнення електронної вакансії в молекулі П680 відбувається за рахунок води. До складу ФС II входить водо-окиснювальний комплекс, що містить в активному центрі іони Мангану в кількості 4 штук. Для утворення однієї молекули кисню потрібно дві молекули води, що дають 4 електрони. Тому процес проводиться в 4 такти, і для його повного здійснення потрібно 4 кванти світла. Комплекс розташований з боку внутрішньотилакоїдного простору, і отримані 4 протони викидаються в нього.
Таким чином, загальний підсумок роботи ФС II — це окислення 2 молекул води за допомогою 4 квантів світла з утворенням 4 протонів у внутрішньотилакоїдному просторі і 2 відновлених пластохинонів в мембрані.
b6f- або b/f-комплекс
b6f-комплекс є насосом, що перекачує протони із строми у внутрішньотилакоїдний простір і створює градієнт їхньої концентрації за рахунок енергії, що виділяється в окисно-відновних реакціях електронно-транспортного ланцюжка. 2 пластохінони забезпечують перекачування 4 протонів. Надалі трансмембранний протонний градієнт (pH строми близький до 8, а внутрішньотилакоїдного простору — до 5) використовується для синтезу АТФ трансмембранним ферментом АТФ-синтазою.
Фотосистема I
Світлозбиральний комплекс I містить приблизно 200 молекул хлорофілу.
У реакційному центрі першої фотосистеми знаходиться димер хлорофілу а з максимумом поглинання при 700 нм (П700). Після збудження квантом світла він відновлює первинний акцептор — хлорофіл а, той — вторинний (вітамін K1), після чого електрон передається на філохінон, від нього на ферредоксин, який і відновлює НАДФ за допомогою ферменту ферредоксин-НАДФ-редуктази. Білок пластоціанін, окиснений в b6f-комплексі, доправляється до реакційного центру першої фотосистеми з боку внутрішньотилакоїдного простору й відновлює окиснений П700.
Циклічний і псевдоциклічний транспорт електрона
Крім повного нециклічного шляху електрона, описаного вище, виявлено циклічний і псевдоциклічний. Суть циклічного шляху полягає в тому, що феридоксин замість НАДФ відновлює пластохінон, який переносить його назад на b6f-комплекс. В результаті утворюється більший протонний градієнт і більше АТФ, але не виникає НАДФН. При псевдоциклічному шляху феридоксин відновлює кисень, який надалі перетворюється на воду і може бути використаний у фотосистемі II. При цьому також не утворюється НАДФН.
Темнова стадія
У темновій стадії за участю АТФ і НАДФН відбувається відновлення CO2 до глюкози. Хоча світло не потрібне для здійснення даного процесу, воно бере участь у його регуляції.
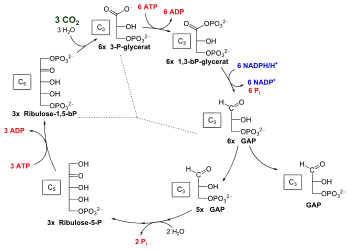
С3-фотосинтез, цикл Кальвіна
Цикл Кальвіна, або відновлювальний пентозо-фосфатний цикл, складається з трьох стадій:
- Карбоксиляція
- Відновлення
- Регенерація акцептора CO2
На першій стадії до рибулозо-1,5-бісфосфату приєднується CO2 під дією ферменту рибулозобісфосфат-карбоксилаза/оксигенази (Rubisco). Цей білок складає основну фракцію білків хлоропласту і, ймовірно, найбільш поширений фермент в природі. В результаті утворюється проміжне нестійке з'єднання, що розпадається на дві молекули 3-фосфогліцеринової кислоти (ФГК). У другій стадії ФГК у два етапи відновлюється. Спочатку вона фосфорилюєтся АТФ під дією фосфорогліцерокінази, потім НАДФН при дії тріозофосфатдегідрогенази, її карбоксильна група окислюється до альдегідної і вона стає вуглеводом (ФГА).
У третій стадії беруть участь 5 молекул ФГА, які через утворення 4-, 5-, 6- і 7-вуглецевих зв'язків об'єднуються в 3 5-вуглецевих рибулезо-1,5-біфосфати, для чого необхідні 3 АТФ. Нарешті, дві ФГА необхідні для синтезу глюкози. Для утворення однієї молекули глюкози потрібно 6 обертів циклу, 6 CO2, 12 НАДФН і 18 АТФ.
С4-фотосинтез, цикл Хетча — Слека — Карпілова
При низькій концентрації розчиненого в стромі CO2 Rubisco каталізує реакцію окислення рибулезо-1,5-біфосфату і його розпад на 3-фосфогліцеринову кислоту і фосфогліколеву кислоту, яка вимушено використовується в процесі фотодихання. Для збільшення концентрації CO2 рослини типу С4 змінили анатомію листка. Цикл Кальвіна у них локалізується в клітинах обкладання провідного пучка, в клітинах мезофілу ж під дією ФЕП-карбоксилази фосфоенолпіруват карбоксилуєтся з утворенням щавлевооцтової кислоти, яка перетворюється на малат або аспартат і направляється в клітини обкладання, де декарбоксилуєтся з утворенням пірувату, що повертається в клітини мезофіла.
С4-фотосинтез практично не супроводжується втратами рибулезо-1,5-біфосфату з циклу Кальвіна, тому ефективніший. Проте він вимагає не 18, а 30 АТФ на синтез 1 молекули глюкози. Це виправдовує себе в тропіках, де жаркий клімат вимагає тримати продихи закритими, що перешкоджає надходженню CO2 в листок, а також при рудеральній життєвій стратегії.
САМ-фотосинтез
У разі CAM-фотосинтезу (Crassulaceae acid metabolism), відбувається розділення асиміляції CO2 і циклу Кальвіна не в просторі, як у С4, а в часі. Вночі у вакуолях клітин по аналогічному описаному вище механізму при відкритих продихах накопичується малат, вдень при закритих продихах йде цикл Кальвіна. Цей механізм дозволяє максимально ощадити воду, проте поступається в ефективності і С4, і С3. Він виправданий при стрестолерантній життєвій стратегії.
Аноксиґенний фотосинтез
Аноксиґенний фотосинтез властивий деяким бактеріям та археям (наприклад, пурпурним, деяким зеленим бактеріям та геліобактеріям тощо). Ці організми не використовують воду як відновник, тому кисень (O2) не є побічним продуктом синтезу. Замість води використовуються як сірководень (H2S) або йони двовалентного заліза (Fe++), унаслідок чого на виході виникають елементарна сірка (S) і тривалентні іони заліза (Fe+++), відповідно, або молекулярний водень (H2). Наприклад, фотосинтез з використанням H2S як відновника проходить такі стадії:
- CO2 + 2 H2S → (CH2O) + 2 S + H2O
Тут першим продуктом фотосинтезу виступає фіктивна хімічна сполука CH2O.
Наприклад, Oscillatoria limnetica, яка живе у гіперсалінному озері Солар-Лейк у районі затоки Ейлат (Ізраїль), може використовувати сульфід як донор електронів, внаслідок чого на поверхні нитки формуються гранули сірки. Взимку у цьому озері відзначаються сильні анаеробні умови, за яких у масі розвивається осциляторія, використовуючи сульфід як донор електронів. Весною, коли води озера насичуються киснем, вона переходить на оксигенний фотосинтез. Таким чином, використання комбінації оксигенного й аноксигенного фотосинтезу, дозволяє осциляторії бути домінуючим фітотрофом цього озера, у якому постійно змінюються аеробні та анаеробні умови.
Окрім осциляторії здатність до факультативного аноксигенного фотосинтезу із використанням сульфіду як донора електронів показана у декотрих штамів Lyngbya, Phormidium, Synechocystis, Prochlorothrix. В ролі донора електронів використовує сірководень Microcoleus chthonoplastes, який переважає у ціанобактеріальних матах й безкисневих умовах у пісках відливно-приливної зони морів.
Здатність до аноксигенного фотосинтезу виявлена у Synechococcus elongatus, який в ролі донора електронів може використовувати тиосульфат. Ця ціанобактерія відкладає сірку не лише у навколишнє середовище, але й у клітинах[3].
Значення фотосинтезу
Фотосинтез є основним джерелом біологічної енергії, фотосинтезуючі автотрофи використовують її для утворення органічних речовин з неорганічних, гетеротрофи існують за рахунок енергії хімічних зв'язків, запасеної автотрофами, вивільняючи її в процесах аеробного та анаеробного дихання. Енергія, отримувана людством при спалюванні викопного палива (вугілля, нафта, природний газ, торф), також є запасеною в процесі фотосинтезу.
Фотосинтез є головним методом залучення неорганічного вуглецю в біологічний цикл. Весь кисень атмосфери біогенного походження є побічним продуктом фотосинтезу. Формування кисневої атмосфери повністю змінило стан земної поверхні, зробило можливою появу дихання, а надалі, після утворення озонового шару, дозволило життю вийти на сушу.
Вивчення фотосинтезу

Перші досліди по фотосинтезу були проведені Джозефом Прістлі в 1770—1780-х роках, коли він звернув увагу на «псування» повітря в герметичній посудині свічкою (повітря переставало бути здатним підтримувати горіння, поміщені в нього тварини задихалися), що горіла, і «виправлення» його рослинами. Прістлі зробив висновок, що рослини виділяють кисень, необхідний для дихання і горіння, проте не відзначив, що для цього рослинам потрібне світло. Це показав незабаром Ян Інгенхауз.
Пізніше було встановлено, що, крім виділення кисню, рослини поглинають вуглекислий газ і за участю води синтезують на світлі органічну речовину. У 1842 Роберт Маєр на підставі закону збереження енергії постулював, що рослини перетворюють енергію сонячного світла в енергію хімічних зв'язків. У 1877 Вільгельм Пфеффер назвав цей процес фотосинтезом.
Хлорофіл був вперше виділений в 1818 році П. Ж. Пелетьє і Жозефом Каванту. Розділити пігменти і вивчити їх окремо вдалося М. С. Цвєту за допомогою створеного ним методу хроматографії. Спектри поглинання хлорофілу були вивчені К. А. Тімірязєвим, він же, розвиваючи положення Маєра, показав, що саме поглинання світла дозволяє підвищити енергію системи, створивши замість слабких зв'язків С-О і О-Н високоенергетичні С-С (до цього вважалося, що у фотосинтезі використовуються жовті промені, що не поглинаються пігментами листка). Зроблено це було завдяки створеному ним методу обліку фотосинтезу за поглинанням CO2, в ході експериментів з освітлення рослини світлом різних довжин хвиль (різного кольору) виявилося, що інтенсивність фотосинтезу збігається зі спектром поглинання хлорофілу.
Окислювально-відновну суть фотосинтезу (як оксигенного, так і аноксигенного) постулював Корнеліс ван Ніль. Це означало, що кисень у фотосинтезі утворюється повністю з води, що експериментально підтвердив 1941 року О. П. Виноградов у дослідах з ізотопною міткою. У 1937 Роберт Хілл встановив, що процес окиснення води (і виділення кисню), а також асиміляції CO2, можна роз'єднати. У 1954—1958 Деніел І. Арнон встановив механізм світлових стадій фотосинтезу, а суть процесу асиміляції CO2 була розкрита Мельвіном Кальвіном з використанням ізотопів вуглецю в кінці 1940-х, за цю роботу в 1961 йому була присуджена Нобелівська премія.
У 1955 була виділена і очищена Rubisco. С4-фотосинтез був описаний Ю. С. Карпиловим в 1960 і М. Д. Хетчемом та К. Р. Слеком в 1966.
Примітки
- Ничипорович А. А. Ничипорович А. А. О Фотосинтезе растений. — М.: Правда, 1948. — 31 с
- Бобось І. М., Завадська О. В. Вплив фотоактивної радіації на врожайність сортів селери в умовах Лісостепу України[недоступне посилання з липня 2019] // Наукові доповіді Національного університету біоресурсів і природокористування України. Електронне наукове фахове видання[недоступне посилання з липня 2019]. — 2012, № 4(33)[недоступне посилання з липня 2019]
- Белякова Г.А. и др. - Ботаника, т.1.
Література
- Мусієнко М. М. Фотосинтез: навч. посібник для студ. вузів, що вивч. дисципліну «Фотосинтез» / М. М. Мусієнко. — К. : Вища шк., 1995. — 247 с.:іл. — ISBN 5-11-004398-1
- Червінський Л. С. Світлокультура рослин — історія виникнення і становлення
- Фотосинтез//Науковотехничний енциклопедичний словник[недоступне посилання з липня 2019]
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
- Физиология растений / под ред. проф. Ермакова И. П. — М.: Академия, 2007
- Молекулярная биология клетки / Альбертис Б., Брей Д. и др. В 3 тт. — М.: Мир, 1994
- Рубин А. Б. Биофизика. В 2 тт. — М.: Изд. Московского университета и Наука, 2004.
Посилання
- Фотосинтез // Словник-довідник з екології : навч.-метод. посіб. / уклад. О. Г. Лановенко, О. О. Остапішина. — Херсон : ПП Вишемирський В. С., 2013. — С. 185.
- Реакції відновлення у біологічних системах#Реакції відновлення у фотосинтезі
- Типи фотосинтезу і організмів-продуцентів[недоступне посилання з липня 2019]
- A collection of photosynthesis pages for all levels from a renowned expert (Govindjee)
- In depth, advanced treatment of photosynthesis, also from Govindjee
- Science Aid: *Реакції Photosynthesis Article appropriate for high school science
- Liverpool John Moores University, Dr.David Wilkinson
- Metabolism, Cellular Respiration and Photosynthesis — The Virtual Library of Biochemistry and Cell Biology
- Overall examination of Photosynthesis at an intermediate level
- Overall Energetics of Photosynthesis
- Photosynthesis Discovery Milestones — experiments and background
- The source of oxygen produced by photosynthesis Interactive animation, a textbook tutorial
