Білок SUMO
Білки МУМО (від Малий Убіквінтин-пов'язаний Модифікатор; англ. Small Ubiquitin-related Modifier, SUMO) — родина маленьких білків, що ковалентно зв'язуються та відділяються від інших білків клітини, змінюючи їх функцію. В середньому їх молярна маса складає 10-12 кДа. Принцип їх дії безпосередньо пов'язаний зі зміною білок-білок або білок-ДНК взаємодій, а також із локалізацією білку в клітині.
Родина білків SUMO
Експресія
Родина білків SUMO представлена в усіх типах клітин. Їх номенклатура наведена у таблиці.
| Людина | Дріжджі | |
|---|---|---|
| SUMO | SUMO1/PIC 1/Ubl1/sentrin1/ GMP1/hSMT3
SUMO2/sentrin3/Smt3a SUMO3/sentrin2/Smt3b |
Smt3 |
| E1 фермент-активатор
субодиниця 1 субодиниця 2 |
SAE1/Aos1/Sua
SAE2/Uba2/ Uba2 |
Aos1
Uba2 |
| Е2 фермент-кон'югатор | Ubc9/UBE2I | Ubc9 |
| Е3 лігаза | Pias1/GBP/DEAD/H box-binding protein 1/ARIP
Piasxb/Miz1/Siz2 RanBP2/Nup358 |
Siz1/UII 1
Siz2/Nfi1 |
У різних сімействах цих білків число членів коливається від одного (наприклад, дріжджі, нематоди і плодова мушка) до кількох тисяч (три у ссавців; вісім у арабідопсис)
Цікаво, що у ссавців білок SUMO1 в основному кон'югований з мембраною, в той час як SUMO2/3 в основному існує у вільній формі.
Механізм регуляції експресії цих білків є недостатньо вивченим, проте встановлено, що вони необхідні для життя більшості організмів.
Структура
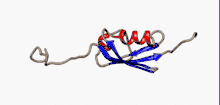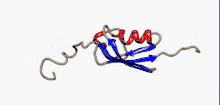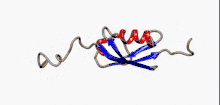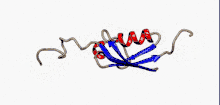
SUMO білки представляють собою невеликі кислі білки, віддалені гомологи убіквітину. Секвенування амінокислотної послідовності дало змогу порівняти ці молекули: білки родини SUMO на 10-20% ідентичні убіквітину. Проте більше подібних ознак вони мають у своїй структурі.
Характерною рисою всіх членів цього сімейства біополімерів, яка відсутня в убіквітину або інших білках, є N-кінцеве гнучне подовження поліпептидного ланцюгу в 10-30 амінокислот. Функція цієї послідовності на сьогодні невідома.
Всі білки SUMO синтезуються у вигляді попередників, які подальшої модифікації - часткового проеолізу C-кінця. Протеолітичне видалення декількох амінокислот, яке досягається специфічними ферментами (SUMO isopeptidases), призводить до оголення C-кінцевого гліцин-гліцинового мотиву. Дозрівання молеули є швидким, проте на даний час немає доказів регулювання цього процесу.
Сумоїлювання
«Сумоїлювання» є посттрансляційною модифікацією, залученою в різні клітинні процеси, такі як ядерний транспорт, регулювання транскрипції, апоптоз, стабільність білків, відповідь на стрес і протікання клітинного циклу.[1]
Протеїни SUMO подібні до убіквітину, а сумоїлювання служить початком ферментного каскаду, аналогічного процесу, що починається з убіквітинювання. На відміну від убіквітину, SUMO не використовується для того, щоб мітити білки, призначені для деградації.
Зрілі білки SUMO утворюються, коли від C-кінця відщеплюються останній чотири амінокислоти. Члени сімейства останній чотири амінокислоти часто мають різнорідні імена; гомолог SUMO1 в дріжджах, наприклад, називається SMT3 (suppressor of mif two 3). Для цього гену повідомлені кілька псевдогенів.
Сумоїлювання як засіб для регулювання функції білка є досить поширеним механізмом: ця модифікація була зареєстрована для більш ніж 70 різних цільових білків, і це число росте, так як продовжуються дослідження цієї родини протеїнів.
Білки-мішені
На основі імунофлуоресцентного аналізу було встановлено, що більшість мішеней для сумоїлювання конститутивно пов'язані з ядерним компартментом. Відповідно до цього, найбільш відомі цільові білки можуть бути пов'язані з ядерними явищами, такими як функціонування хроматину, репарація ДНК, транскрипція, або цитоплазматичний транспорт (серед них є гістондезацетилази, топоізомерази, тимін-ДНК-глікозилаза, PCNA, p53, PML, фактори теплового шоку, рецепторів стероїдних гормонів, IκBα , RANGAP1 і багато інших).
Іншими прикладами є прогресування шляхом мітозу (мітоз дріжджових мутантів, в яких дефектні білки SUMO — сполучені або декон'юговані з дріжджовим септином, топоізомеразою II і Pds5) або інфікування вірусами (приклади для вірусного організму мішенями SUMO є: для цитомегаловірусу ранні білки IE1 і IE2, аденовірусу 5 Е1В-55 кДа, бичачий папіломавірус E1).
Наслідки модифікації
Подібно до фосфорилювання, сумоїлювання, має багато різних функціональних наслідків, які залежать від специфіки білку-мішені. З огляду на розмір SUMO, можна припустити, що приєднання може призвести до маскування сайтів зв'язування, генерації неправильного зв'язування, конформаційних змін в модифікованому білку.
Так, були зареєстровані зміни в білок-білок або білок-ДНК взаємодіях, зміни у внутрішньоклітинній локалізації, підвищенні стабільності білку за рахунок антагонізму із убіквітин /протеасомною деградацією, а також зміни в ферментативній активності протеїнів.
Посилання
- Hay RT (Apr 2005). SUMO: a history of modification. Mol. Cell 18 (1): 1–12. PMID 15808504. doi:10.1016/j.molcel.2005.03.012.[недоступне посилання з березня 2019]