Вухо
Ву́хо — це орган у хребетних тварин (вкл. й людину), що працює для сприйняття звуку та підтримання рівноваги у просторі. Людське вухо здатне сприйняти звукові хвилі з довжиною від 1,6 см до 20 м, що відповідає 16 — 20 000 Гц (коливань в секунду).

Анатомія вуха людини
Вухо людини та інших ссавців складається з трьох частин: зовнішнє, середнє та внутрішнє вухо. Зовнішнє і середнє вухо побудовані відносно просто, їхня функція полягає у проведенні і підсиленні звукових коливань. Внутрішнє вухо найскладніше за будовою, крім сприйняття звуків також забезпечує рівновагу і відчуття положення тіла в просторі[1].
Зовнішнє вухо
- Основна стаття: Зовнішнє вухо
Зовнішнє вухо складається із вушної раковини (лат. auricula) та зовнішнього слухового проходу. Вушна раковина (саме цю частину органа слуху більшість людей називають вухом) оточує зовнішній слуховий прохід. Вона складається із еластичного хряща, покритого шкірою. Деякі частини вушної мушлі, зокрема мочка, позбавлені хрящової тканини. Функція вушної раковини полягає у тому, що вона спрямовує звук у зовнішній слуховий прохід[2].
Дарвінів горбик

У деяких людей (близько 10 %) на верхній частині завитки вушної раковини наявний невеликий виступ, який називають Дарвінів горбок (лат. tuberculum auriculae). Він вважається рудиментом загостреної верхівки вуха у деяких приматів та інших ссавців. Свою назву Дарвінів горбик отримав за те, що у своїй праці «Походження людини і статевий добір» Дарвін згадує його як приклад рудиментарного органа[3].
Зовнішній слуховий прохід
Зовнішній слуховий прохід — це коротка зігнута трубка (2,5 см довжиною і 0,6 см у діаметрі), що веде до барабанної перетинки. Поблизу вушної раковини зовнішній слуховий прохід підтримується еластичним хрящем, решта його проходить у скроневій кістці. Весь канал вистелений шкірою із коротким волоссям, потрібним для того, щоб затримувати різні сторонні об'єкти, наприклад пил і дрібних комах. Також у шкірі зовнішнього слухового проходу наявні сальні залози і модифіковані апокринні потові залози, які ще називають церумінозними, вони виділяють жовто-коричневу воскоподібну вушну сірку. Сірка досить липка, тому вона, як і волосся, може перешкоджати просуванню сторонніх об'єктів по зовнішньому слуховому проходу, крім того вона є репелентом для комах і пригнічує ріст мікроорганізмів[2]. Сірка складається із насичених та ненасичених довголанцюгових жирних кислот, спиртів, сквалену (приблизно 12—20 % складу сірки), та холестеролу (6—9 %). Разом із сіркою із зовнішнього вуха виводяться частинки рогового шару шкіри, який постійно злущується, тому вона може містити багато білків кератинів[4].
У деяких людей вуха очищуються від сірки природним шляхом: вона засихає і випадає із зовнішнього слухового проходу, цьому сприяють рухи щелепи, наприклад при жуванні або розмові. В інших людей сірка може накопичуватись, ущільнюватись і перешкоджати слуху.
Барабанна перетинка
На межі між зовнішнім та середнім вухом розташована барабанна перетинка (лат. membrana tympani). Це тонка прозора мембрана з волокнистої сполучної тканини, із зовнішнього боку вкрита шкірою, а із внутрішнього — слизовою оболонкою. По периферії перетинки волокна сполучної тканини розташовані переважно по колу, а в центрі — радіально. Барабанна перетинка має форму сплощеного конуса, верхівка якого спрямована у порожнину середнього вуха. Вона прикріплена волокнисто-хрящовим кільцем до барабанної борозни. Барабанна перетинка коливається під впливом звукових хвиль, і передає ці коливання на слухові кісточки середнього вуха[2].
Середнє вухо
- Основна стаття: Середнє вухо
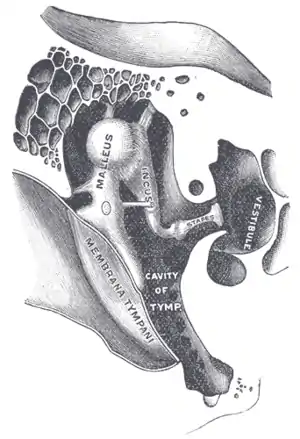
Середнє вухо (барабанна порожнина) — це невелика заповнена повітрям і вистелена слизовою оболонкою порожнина у скроневій кістці. З одного боку вона обмежена барабанною перетинкою, а з іншого — кістковою стінкою із двома отворами: овальним та круглим вікном внутрішнього вуха. Верхня частина середнього вуха аркоподібно підіймається вгору і утворює надбарабанний закуток. На медіальній стінці середнього вуха розміщена соскоподібна печера, що дозволяє йому контактувати із соскоподібними комірками скроневої кістки[5].
Нижня частина барабанної порожнини містить отвір слухової (фаринготимпанальної або євстахієвої) труби, що направалена вниз і сполучає порожнину середнього вуха із носоглоткою. Більшість часу слухова труба сплощена і закрита, відкривається вона тільки під час зівання або ковтання для того щоб зрівняти тиск у середньому вусі із тиском у зовнішньому середовищі. Це важливо для того, щоб барабанна перетинка могла нормально коливатись. Якщо існує різниця тисків зовні та всередні барабанної порожнини, це супроводжується відчуттям «закладених вух», яке виникає при різкій зміні висоти, наприклад під час злітання чи приземлення літака.
У порожнині середнього вуха розташовані слухові кісточки:
Руків'я молоточка приєднане до барабанної перетинки, а основа стремінця до овального вікна внутрішнього вуха. Слухові кісточки підтримуються зв'язками, що відходять від стінок барабанної порожнини, між собою вони сполучені синовіальними суглобами. Основна роль слухових кісточок полягає у передачі коливань барабанної перетинки на перетинку овального вікна. Оскільки барабанна перетинка має у 22 рази більшу площу ніж перетинка овального вікна, при цьому відбувається значне підсилення коливань.
Також у барабанній порожнині розміщені два маленькі скелетні м'язи:
- м'яз-натягувач барабанної перетинки, який починається від стінки слухової труби і закінчується на молоточку
- стремінцевий м'яз, натягнений між задньою стінкою барабанної порожнини і стремінцем.
Якщо вухо подразнююється дуже голосним звуком, ці м'язи рефлекторно скорочуються і зменшуюють коливання овального вікна внутрішнього вуха, таким чином запобігаючи ушкодженню слухових рецепторів[2].
Внутрішнє вухо
- Основна стаття: Внутрішнє вухо
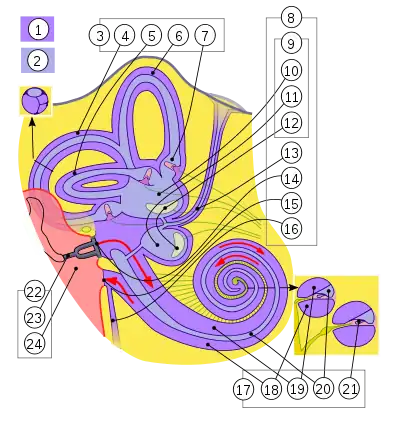
Внутрішнє вухо через свою складну форму також називається лабіринт. Воно лежить глибоко у скроневій кістці позаду очних впадин. Внутрішнє вухо складається із двох основних частин:
- Кістковий лабіринт — це система звивистих каналів у скроневій кістці, він заповнений перилімфою — рідиною, аналогічною за складом до спинномозкової рідини, вони можуть перетікати одна в одну через водопровід завитки;
- Перетинчастий лабіринт — це серія мебранних мішечків та проток, що містяться всередині кісткового лабіринту. Він заповнений ендолімфою, що за хімічним складом близька до вунтрішньо клітинної рідини багатої на іони K+. В утворенні ендолімфи бере участь ендолімфітичний мішок, що сполучений із рештою перетинчастого лабіринту ендолімфатичою протокою[2][5].
До складу внутрішнього вуха входять три основні відділи: присінок (переддвер'я), півколові канали та завитка.
Присінок
Присінок або переддвер'я (лат. vestibulum) — це центральна яйцеподібна порожнина кісткового лабіринту. Він лежить позаду завитки, і попереду від півколових каналів. У перилімфі присінка розташовані два з'єднані протокою мішечки перетинчастого лабіринту:
- овальний або просто мішечок (лат. sacculus), сполучений із завиткою;
- круглий або маточка (лат. utriculus) сполучена із півколовими каналами.
У мішечку та маточці розташовані рецепторні органи рівноваги, що називаються плямами (лат. maculus), вони подразнюються у відповідь на зміну положення голови[2].
Півколові канали
Півколові канали розміщені позаду присінка, кожен канал становить близько двох третин кола. У людини наявні три півколові канали розташовані у взаємноперпендикулярних полощинах: передній, задній і латеральний. Передній і задній розташовані вертикально під прямим кутом одне до одного, а латеральний — горизонтально. Кожен канал має на одному із кінців розширення, що називається ампула, в ампулах знаходяться ампульні гребінці, які забезпечують відчуття кутового прискорення, їх рецептори подразнюються при обертанні голови[2].
Завитка
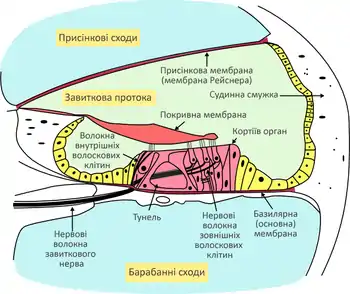
Завитка (лат. cochlea) — конічна закручена комірка у кістці, розміром приблизно із половинку горошини. Завитка робить приблизно 2,75 оберту, в ній розміщена завиткова протока, де міститься спіральний або кортіїв орган — рецепторний орган слуху. Завиткова протока разом із спіральною кістковою пластинкою поділяють порожнину кісткової завитки на три відсіки або сходи:
- присінкові сходи (лат. scala vestibuli), розміщені над завитковою протокою, сполучені із присінком і впираються в овальне вікно;
- середні сходи — це сама завиткова протока;
- барабанні сходи (лат. scala tympani) розміщені під завитковою протокою і закінчуються круглим вікном.
Завиткова протока як частина перетинчастого лабіринту заповнена ендолімфою. Присінкові і барабанні сходи, як частина кісткового лабіринту — перилімфою, на верхівці завитки вони сполучені між собою через отвір — гелікотрему.
Верхню стінку завиткової протоки утворює присінкова перетинка або мембрана (мембрана Рейснера), що відмежовує її від присінкових сходів. Зовнішній край присінкової мембрани складається із особливо багатої на кровоносні судини слизової оболонки, що називається судинна смужка (лат. stria vascularis), вона бере участь в утворенні ендолімфи. Нижня стінка завиткової протоки утворена спіральною кістковою пластинкою та гнучкою волокнистою базилярною або основною мембраною, на якій розміщений Кортіїв орган. Базилярна мембрана відіграє дуже важливу роль у сприйнятті звуків, вона вузька і товста поблизу овального вікна і стає ширшою і тоншою до верхівки завитки[2].
Будова Кортієвого (спірального) органа

Кортіїв орган розташований на поверхні базилярної мембрани, він складається із підтримуючих клітин та слухових рецепторів — волоскових клітин завитки, зверху ці клітини вкриті покривною мембраною. Волоскові клітини розташовються чотирма рядами: один ряд внутрішніх волоскових клітин і три ряди зовнішніх. До основи волоскових клітин підходять чутливі нервові закінчення завиткового нерва (гілки присінково-завтикового або 8-ї пари черепномозкових нервів). На верхівці волоскові клітини мають велику кількість довгих мікроворсинок (стереоцілії) та одну війку (кінетоціль). Всередині мікроворсинки укріплені актиновими філаментами, поблизу верхівок вони сполучені між осбою спеціальними білковими місточками. Стереоцілії та кінотоцілії виступають у ендолімфу, багату на іони калію, найдовші із них впираються у покривну мембрану[2].
Фізіологія сприйняття звуку
Відчуття звуку виникає у людини при стимуляції слухової кори скроневої частки головного мозку. Проте до того як це станеться звукові хвилі повинні пройти шлях через зовнішній слуховий прохід, барабанну перетинку, кісточки середнього вуха, перетинку овального вікна, перелімфу завитки, передатись на базилярну мембрану, сприйнятись волосковими клітинами Кортієвого органа, які передають збудження на чутливі нервові закінчення присінково-завиткового нерва, по якому імпульси і надходять до головного мозку.
Передача і підсилення звукового сигналу у вусі
Звукові хвилі через зовнішній слуховий отвір надходять до барабанної перетинки і спричинюють її коливання із відповідною частотою та амплітудою. Чим більша інтенсивність (гучність) звуку, тим більша амплітуда коливань перетинки. Рух барабанної перетинки передається на слухові кісточки, що діють як важелі і розгойдують перетинку овального вікна внутрішнього вуха. Оскільки площа цієї перетинки у 20-22 рази менша за барабанну, то амплітуда коливань зростає тут у відповідну кількість разів, тобто відбувається підсилення сигналу.
Коливаючись вперед-назад із певною частотою, овальне вікно внутрішнього вуха спричиняє у присніковому каналі аналогічний рух перелімфи, що поширюється від основи до верхівки завитки. Звук дуже низької частоти (нижче 16 Гц) створює у рідині хвилі тиску, що проходять всесь шлях від овального вікна, вверх по присінкових сходах, через гелікотерму до барабанних сходів і нарешті передаються на кругле вікно внутрішнього вуха. Такі звуки не подразнюють кортієвого органа і перебувають поза межами чутливості людського вуха (інфразвук). Звукові хвилі більшої частоти створюють хвилі тиску, які можуть «зрізати» шлях через вестибулярний канал до перилімфи барабанних сходів. У такому разі вони спричинюють коливання базилярної мембрани, внаслідок чого покривна мембрана зміщується, ритмічно подразнюючи волоскові клітини.
Сприйняття звукових коливань волосковими клітинами Кортієвого органа
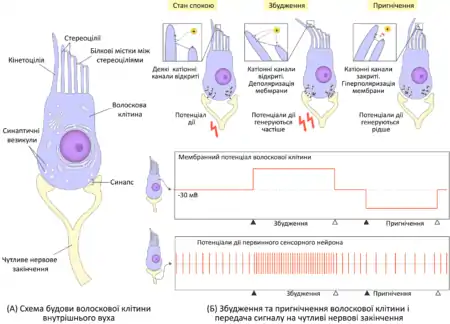
Механорецепторами, що сприймають звук у внутрішньому вусі є волоскові клітини. Коли під впливом коливань базилярної мембрани покривна мембрана зміщується, вона згинає довгі стереоцілі волоскових клітин у сторону до кінетоцілі. При цьому білкові фібрили які сполучають їх з короткими волосками розтягаються і відкривають катіонні (калієві і кальцієві) канали, через які всередину клітини рухаються іони K+ і Ca2+ і спричинюють деполяризацію її мембрани. Таким чином у волоскових клітинах виникає рецепторний потенціал, що викликає вивільнення неймедіатора (глутамату) у синаптичну щілину. Збудження передається на чутливі нервові закінчення і вони генерують потенціали дії із більшою частотою. Коли покривна мембрана завиткової протоки зміщується в протилежну сторону, стереоцілі волоскових клітин відхиляються від кінетоцілі, при цьому катіонні канали закриваються і відбувається реполяризація мембрани, відповідно синаптична передача блокується і чутливі нервові закінчення генерують нервові імпульси із меншою частотою. Регулярна зміна частоти нервових імпульсів і сприймається головним мозком як відчуття звуку.
Волоскові клітини відрізняються за своїми функціями. Внутрішніх волоскових клітин розміщених в один ряд в тричі менше ніж зовнішніх, але вони відповідають за передачу 90—95 % інформації до мозку. Зовнішні волоскові клітини, розміщені у три ряди, навпаки, отримують еферентні сигнали від мозку і беруть участь у підсиленні або послабленні коливань базилярної мембрани. Під дією ритмічної деполяризації і реполяризації зовнішні волосокові клітини починають ритмічно скорочувтись і розтягуватись, змінюючи натяг базилярної мембрани, таким чином вони збільшують її коливання у певному місці і підсилюють точність сприйняття звуку. Вважається, що саме внаслідок їхнього руху виникає отоакустична емісія, тобто із самого вуха виходять звуки. Це явище використовують для виявлення вад слуху у новонароджених. Під впливом дуже гучних звуків мозок надсилає до зовнішніх волоскових клітин сигнал, у відповідь на що вони перестають розгойдуватись, тому коливання базилярної мембрани поширюються на більшу відстань. Це потрібно для того, щоб захистити волоскові клітини від ушкодження.
Сприйняття характеристик звуку
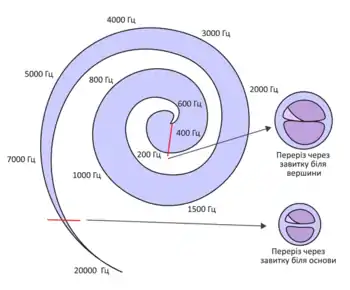
Людина здатна сприймати частоту, гучність та напрям надходження звукової хвилі, кожна із цих характеристик оцінюється головним мозком на основі різних механізмів:
- Сприйняття частоти звуку. Інформація про висоту звуку кодується ще у внутрішньому вусі, хвилі різної частоти сприймаються в залежності від того, в якій самі частині завитки вони викликають збудження волоскових клітин. Базилярна мембрана містить велику кількість волокон, натягнутих поперек неї, при чому вони не однакові по всій її довжині. Біля основи завитки ці волокна коротші і тугіші, а сама мембрана товстіша, біля верхівки волокна коротші і більш провислі. Через це звуки високої частоти викликають максимальне коливання базилярної мембрани відразу ж біля овального вікна, а низької частоти — ближче до гелікотерми. Отже якщо до мозку надходять нервові імпульси від волоскових клітин нижньої частини завитки, він інтерпретує їх як низький звук, із верхньої — як високий. Звук, що складається із кількох тонів одночасно активує різні групи волоскових клітин.
- Сприйняття інтенсивності звуку. Гучніший звук викликає коливання барабанної перетинки, слухових кісточок, овального вікна та перилімфи із більшою амплітудою, це у свою чергу викликає сильніше зміщення базилярної мембрани і більше відхилення волосків волоскових клітин, через що виникає рецепторинй потенціал більшої амплітуди і в синаптичну щілину вивільняється більше нейромедіаторів. В такому випадку нервові закінчення завиткового нерва частіше генерують потенціал дії. Мозок інтерпретує це як більшу гучність.
- Сприйняття напрямку звуку. Відчуття напрямку звуку здійснюється на основі порівняння інтенсивності та часу надходження звукових сигналів до кожного із двох вух. У визначенні напрямку звуку беруть участь ядра стовбура головного мозку.
Еволюція елементів вуха
Внутрішнє вухо, як орган слуху та рівноваги, виник ще у перших хребетинх і з того часу зазнав багато ускладнень у процесі еволюції. Крім того апарат слуху поступово доповнювався середнім вухом (вперше з'являється в амфібій) та зовнішнім, наявного у птахів і ссавців.
Еволюція внутрішнього вуха
Внутрішнє вухо (лабіринт) у хребетних тварин виникло як орган рівноваги. Воно складалось із присінка, до складу якого входять круглий і овальний мішечки (круглий мішечок ще називають маточка[6]), а також півколові канали. У міксин наявна тільки одна пара півколових каналів, у міног — дві, у всіх інших хребетних, тобто у щелепоротих (від хрящових і променеперих до птахів і ссавців) — три[7].
У круглоротих основа овального мішечка утворює невелику кишеню, яка називається лагена, і одночасно із забезпеченням рівноваги тіла бере участь у сприйнятті звукових сигналів[8]. В еволюції хребетних лагена перетворилась в орган слуху амфібій. У рептилій вона має дещо більший розмір, і розділена на три канали (як і завитка ссавців). У птахів лагена ще більш витягнена, що дозволяє їм краще чути. Для ссавців характерна найскладніша будова внутрішнього вуха, лагена перетворюється на закручену завитку.
Еволюція слухових кісточок середнього вуха
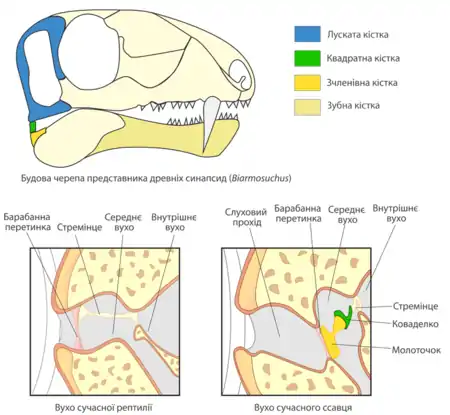
У процесі формування наземних хребетних, зокрема чотириногих (Tetrapoda), відбулися значні зміни в будові вісцерального скелета, які врешті завершилися формуванням слухових кісточок: спочатку стремінця (в амфібій, плазунів та птахів), а потім ще двох — коваделка та молоточка — у ссавців.
Формування стремінця забезпечено вивільненням гіомандибулярної кістки з системи підвіски щелеп, що відбулося ще на стадії формування групи хоанових, або легеневодишних хребетних (Choanata). Ця кісточка, топографічно пов'язана зі спіракулюмом («бризкальцем»), яка надалі стала порожниною середнього вуха, взяла на себе функцію передачі коливань з покривних утворів до власне вуха.
Ця кісточка, що має назву стремінце, або стовпчик, є гомологічною у всіх тетрапод — амфібій, плазунів, птахів і ссавців. Вона має паличкоподібну форму з гострішим внутрішнім кінцем. Гомологічна кістка у риб (гімандибулярна) виконувала функцію опори щелеп.
Формування системи з трьох кісточок середнього вуха у ссавців є одним із найкраще задокументованих за викопними рештками. Їхня поява так само пов'язана із втратою кістками вісцерального скелета своїх первинних функцій. У ссавців це відбулося у зв'язку з тим, що формування мандибули (нижньої щелепи) відбувалося за рахунок лише однієї — зубної — кістки. Інші кістки, що брали участь у формуванні мандибули в ранніх амніот, аналогічно до гіомандибулярної не зникли, а відійшли у область середнього вуха і сформували дві нові слухові кісточки:
- квадратна кістка верхньої щелепи рептилій перетворилась у коваделко,
- зчленівна кістка нижньої щелепи — у молоточок.
Гомологія слухових кісточок ссавців і кісток щелеп рептилій добре досліджена на матеріалах з викопними рештками та даних ембріології ссавців[7].
Особливості будови вуха різних груп хребетних тварин
| Група хребетних | Особливості будови вуха |
| Круглороті | Наявне тільки внутрішнє вухо, складається із присінка та півколових каналів (у міксин — одна пара, у міног — дві). Функцію слуху виконує невеликий виріст овального мішечка — лагена. |
| Хрящові і кісткові риби | Внутрішнє вухо доповнене третім півколовим каналом. Овальний мішечок, круглий мішечок та лагена містять статоліти, вільно приєднані двома мембранами до стінок присінка, таким чином, що вони можуть вібрувати. Коливаючись статоліти подразнюють сенсорний епітелій. У риб групи Ostariophysi слух особливо гострий, частково це забезпечується тим, що в них є спеціальні кісточки (апарат Вебера), що розвиваються із хребців[9]. Апарат Вебера сполучає плавальний міхур із стінкою внутрішнього вуха і передає на нього коливання[8]. |
Амфібії
 Litoria caerulea. На фотографії видно барабанну перетинку |
У земноводних з'являється середнє вухо, що являє собою порожнину, зовнішня сторона якої затягнута барабанною перетинкою. В середньому вусі розміщена паличкоподібна слухова кісточка — стремінце, що одним кінцем упирається в овальне вікно внутрішнього вуха, а другим — у барабанну перетинку. Середнє вухо з'єднане із ротоглоткою євстахієвою трубою. У хвостатих середнє вухо відсутнє[8].
Лагена більша ніж у риб і частково вкрита покривною (текторальною) мембраною. Ця структура зазвичай чутлива до низькочастотних звуків (не більше 4000 Гц). Наприклад велика зелена жаба чує звуки від 100 до 200 Гц, тобто із частотою, що відповідає крикам самців[7]. |
Рептилії
 Dracaena guianensis. На фотографії видно барабанну перетинку |
Слух розвинений добре. Вперше з'являється стуркутра схожа на завитку: у лагені наявні три канали, дно лагени формує базилярну мембрану. У всіх рептилій крім змій є середнє вухо. У змій стремінце приєднане до квадратної кістки щелепи, тому вони не переважно погано чують звуки у повітрі, але добре вловлюють коливання землі[8]. |
Птахи
 Казуар звичайний. На фотографії видно зовнішній слуховий прохід |
Вухо має три відділи: внутрішнє, середнє і зовнішнє вухо, останнє представлене зовнішнім слуховим проходом. У внутрішньому вусі розміщена завитка, вона коротша ніж у ссавців і не закручена. Більшість птахів можуть чути приблизно у тому ж діапазоні частот, що й людина. Проте ссавці такого ж розміру здатні сприймати більш високочастотні звуки. Птахи добре відрізняють частоти звуків, і можуть встановлювати місце, звідки надходить звук[7]. |
| Ссавці | Особливістю будови вуха ссавців є наявність вушної мушлі, трьох слухових кісточок у середньому вусі і закручена завитка. В залежності від способу життя вушні мушлі різних ссавців відрізняються за будовою. У більшості звірів, наявні спеціальні м'язи, які дозволяють повертати вуха. В інших ссавців, як і у людини, рухливість вушної мушлі різко обмежена. У багатьох форм, які ведуть водний (китоподібні, тюлені, моржі) або підземний спосіб життя (кріт), вушні раковини редукуються.
Будова внутрішнього вуха у різних ссавців також дещо відрізняється. Наприклад, кількість поворотів коливається від чверті у качкодзьоба до чотирьох у свині і морської свинки. У кита — півтора повороту, у коня — 2, у людини — 2,75, у кота — 3[8]. Особливо тонкий слух мають звірі, активність яких найбільша в нічний час[10]. Верхня межа чутливості у псів — 45 кГц, у котів — 50 кГц. Деякі ссавці, зокрема кажани і китоподібні, мають здатність до ехолокації, верхня межа чутливості їхнього вуха сягає 100 кГц[8]. |


.jpg.webp)
Джерела
- Campbell NA, Reece JB (2008). Biology (вид. 8th). Benjamin Cammings. ISBN 978-0321543257.
- Marieb EN, Hoehn K (2006). Human Anatomy & Physiology (вид. 7th). Benjamin Cummings. ISBN 978-0805359091.
- [ http://www.britannica.com/EBchecked/topic/152008/Darwins-tubercle Encyclopædia Britannica]
- >Guest JF, Greener MJ, Robinson AC, Smith AF (August 2004). Impacted cerumen: composition, production, epidemiology and management. QJM 97 (8): 477–88. PMID 15256605. doi:10.1093/qjmed/hch082.
- Неттер Ф. Атлас анатомії людини/ Під ред. Проф. Ю.Б.Чайковського/ Наук пер. з англ. к.м.н. Цегельського А.А. – Львів:Наутілус, 2004. -592с. ISBN 966-95745-8-7
- Тейлор Д., Грин Н., Стаут У. Биология: В 3-х т. Т.2: Пер.с англ./Под ред. Р.Сопера — 3-е изд. — М.:Мир,2004 —436с.
- Hickman CP Jr., Roberts LS, Larson A (2001). Integrated principles of zoology (вид. 11th). McGraw-Hill Higher Education. ISBN 0–07–290961–7.
- Prosser CL, Bishop DV, Brown FA Jr., Jahn TL, Wulf VJ (1950). Comparative animal physiology. W.B.Saunders Company.
- Encyclopædia Britannica
- Біологія:Навч. посіб. / А.О.Слюсарєв, О.В.Самсонов, В.М. Мухін та ін.; За ред. та пер. з рос. В.О.Мотузного. – 3-тє вид., випр. і допов. – К.:Вища шк., 2002. – 622с.: іл.
Посилання
- Тест слуху на відчуття різних частот (англ.)
- Відео про сприйняття звуку від Brandon Pletsch (англ.)
- Відео про сприйняття звуку від MED-EL (англ.)
- Інтерактивна програма для вивчення будови вуха (англ.)
- Фотографії елементів вуха зроблені за допомогою скануючого електронного мікроскопа