Глікогеноліз
Глікогено́ліз — внутрішньоклітинний процес розщеплення глікогену до глюкозо-6-фосфату для його подальшого використання в процесах енергетичного обміну. Найбільш інтенсивно відбувається в основних місцях зберігання глікогену: скелетних м'язах, де більшість утвореного глюкозо-6-фосфату вступає у гліколіз для забезпечення енергетичних потреб, і в печінці, де глюкозо-6-фосфат дефосфорилюється і виділяється у кров для підтримання стабільного рівня глюкози.
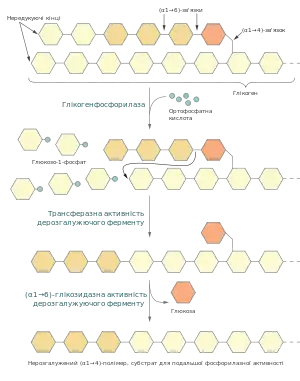
Для проходження глікогенезу необхідна активність трьох ферментів: глікогенфосфорилази, біфункціонального дерозгалужуючого ферменту та фосфоглюкомутази. Процес контролюється на кількох рівнях, його стимулюють гормони глюкагон та адреналін.
Механізм
На відміну від процесу перетравлення глікогену внутрішньоклітинний глікогеноліз відбувається шляхом фосфоролізу, а не гідролізу. Тобто глікозидні зв'язки полімеру атакуються не водою, а фосфатною кислотою. Відповідно продуктом реакції є не вільна глюкоза, а глюкозо-1-фосфат. Утворення естеру дозволяє зберегти велику частину енергії глікозидних зв'язків і для включення моносахариду у метаболічні шляхи, такі як гліколіз та пентозофосфатний шлях не потрібно їх додатково фосфорилювати, на що витрачалась би молекула АТФ. Крім того утворення глюкозо-1-фосфату у м'язах має ще одну перевагу: ця сполука негативно заряджена за фізіологічних умов і для неї, на відміну від глюкози, нема переносників у плазматичній мембрані, тому вона не може покидати клітину[1][2].
Глікогенфосфорилаза
Процес гілкогенолізу починається із фосфоролізу (α1→4)-глікозидних зв'язків із нередукуючого кінця, що каталізується глікогенфосфорилазою. Коферментом у реакції є пірідоксаль-5'-фосфат, ковалентно приєднаний до ферменту через шифову основу із лізином у положенні 679[3]. Фосфатна група цієї молекули функціонує як загальний кислотний каталізатор, що стимулює атаку глікозидного зв'язку ортофосфатною кислотою[4].

In vitro реакція каталізована глікогенфосфорилазою є оборотною, значення стандартної зміни вільної енергії ΔG0' є невеликим, через те, що глікозидний зв'язок та естерний, на який він замінюється, мають приблизно однаковий запас енергії. Проте враховуючи, що за pH 6,8 рівноважне співвідношення [Фн]/[глюкозо-1-фосфат] становить близько 3,6, а в цитоплазмі клітини воно зазвичай перевищує 100, рівновага реакції сильно зміщена у сторону розщеплення глікогену (утворення глюкозо-1-фосфату)[1].
Глікоген-зв'язуюча ділянка фосфорилази має форму тріщини, в якій можуть поміститись 4—5 залишків глюкози. Але вона надто вузька для того, щоб захоплювати місця галуження[3]. Тому дія глікогенфосфорилази припиняється у точці віддаленій на чотири залишки глюкози від (α1→6)-зв'язку[4]. Аткивність цього ферменту є предметом складних механізмів регулювання, як алостеричного так і шляхом ковалентної модифікації[5].
Дерозгалужуючий фермент
Якщо б гліконефосфорилаза діяла сама, то розщеплення глікогену завершувалось би після вивільнення близько шести залишків глюкози на гілку. Для того, щоб деполімеризувати цілу молекулу потрібні дві додаткові ферментативні активності: трансферазна та α-1,6-глікозидазна, у клітинах еукаріот обидвома активностями володіє один так званий дерозгалужуючий фермент. Спершу він каталізує перенесення фрагменту із трьох глюкозних залишків із зовнішньої гілки на внутрішню. Після цього залишається один глюкозний мономер приєднаний (α1→6)-глікозидним зв'язком, який гідролізується завдяки α-1,6-глікозидазній активності[6][4]:

Оскільки відбувається реакція гідролізу, а не фосфоролізу, то продуктом є глюкоза, а не глюкозо-1-фосфат. Для подальшого використання у гліколізі глюкоза фосфорилюється гексокіназою[6].
У процесі глікогенезу розгалуження відбувається іншим чином: трансферазна реакція, каталізована розгалужуючим ферментом (аміло-(1,4→1,6)-трансглікозилазою), полягає у гідролізі (α1→4)-зв'язку та утворенні утворенні (α1→6)-зв'язку. Під час глікогенолізу зворотна реакція не відбувається (трансферазна активність дерозгалужуючого ферменту полягає у гідролізі одного та утворенні іншого (α1→4)-зв'язку). Це пояснюється з точки зору енергетики процесу: зміна вільної енергії під час гідролізу (α1→4)-зв'язку становить −15,5 кДж/моль, а для гідролізу (α1→6)-зв'язку — −7,1 кДж/моль. Через це розщеплення (α1→4)-зв'язку може забезпечувати синтез (α1→6)-зв'язку, але не навпаки[7].
Утворений лінійний фрагмент молекули глікогену знову стає субстратом для глікогенфосфорилази, що продовжує розщеплення до наступного місця галуження, де у дію повторно вступає дерозгалужуючий фермент.
Фосфоглюкомутаза
Для того, щоб вступити у метаболічні шляхи клітини, глюкозо-1-фосфат повинен бути перетворений у глюкозо-6-фосфат. Це здійснюється завдяки фосфоглюкомутазі, яка окрім того бере участь у метаболізмі галактози. Реакція відбувається шляхом обміну фосфатними групами між ферментом та субстратом[8]:

Для того щоб фосфоглюкомутаза могла каталізувати реакцію, залишок серину в активному центрі повинен бути фосфорильованим. Іноді під час реакції відбувається дисоціація глюкозо-1,6-бісфосфату, внаслідок чого фермент залишається в інактивованому стані. Для забезпечення його реактивації в клітині завжди підтримується деякий рівень глюкозо-1,6-бісфосфату, який синтезується ферментом фосфоглюкокіназою[9].
Використання глюкозо-6-фосфату
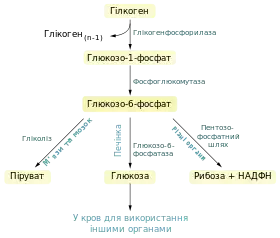
Продукт глікогенолізу глюкозо-6-фосфат може перетворюватись трьома шляхами: він може бути субстратом для гліолізу (переважно у м'язах та мозку), перетворюватись у вільну глюкозу і поступати в кров (у печінці, і, меншою мірою, в нирках) або перетворюватись у пентозофосфатному шляху із утворенням рибози та НАДФH.
Перетворення у вільну глюкозу
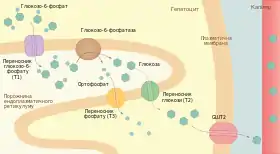
Зі всіх тканин організму тільки печінка та нирки містять фермент глюкозо-6-фосфатазу, він необхідний для поповнення рівня глюкози в крові у період між прийомами їжі або інтенсивного фізичного навантаження. Глюкозо-6-фосфатаза — інтегральний фермент мембрани ендоплазматичного ретикулуму із активним центром повернутим в його порожнину. Глюкозо-6-фосфат, що утворюється у цитоплазмі в процесі глікогенолізу, транспортується спеціальним переносником (T1) в ЕР, де відбувається його гідроліз. Продукти — глюкоза та фосфат — транспортуються назад у цитозоль іншими білками — T2 та T3, а звідти за участі переносника GLUT2 глюкоза виділяється у кров[4][8].
Таким чином у гепатоцитах та клітинах нирок два шляхи використання глюкозо-6-фосфату — гліколітичний та перетворення у вільну глюкозу — розділені просторово[4].
Регуляція глікогенолізу
Лімітуючу стадію глікогенолізу каталізує глікогенфосфорилаза, відповідно, саме вона і є ключовим регуляторним ферментом цього шляху. Каталітична активність глікогенфосфорилази контролюється на двох рівнях: вона приводиться у відповідність із потребами окремої клітини в конкретний момент часу завдяки алостеричній регуляції, окрім того існує «централізована» регуляція завдяки гормонам, таким як адреналін, глюкагон та інсулін, що викликають фосфорилювання або дефосфорилювання ферменту і, відповідно, його активацію або деактивацію[10].
Печінка та скелетні м'язи мають свої особливості регуляції глікогенолізу завдяки наявності різних ізоформ глікогенфосфорилази. Відмінності пов'язані різницею у призначенні розщеплення глікогену в цих органах: в той час як м'язи використовують глюкозу тільки для власних потреб, печінка забезпечує нею весь організм[10].
Активація глікогенолізу
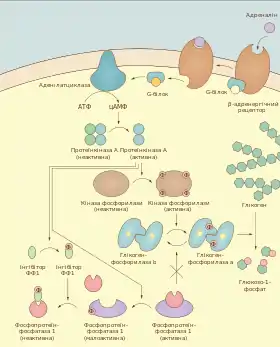
У скелетних м'язах активність глікогенфосфорилази безпосередньо залежить від енергетичного статусу клітини. Аденозинмонофосфат, високі концентрації якого свідчать про вичерпування запасів АТФ, діє як алостеричний активатор цього ферменту. Печінкова ізоформа не чутлива до АМФ[11][12].
Як у м'язах, так і в печінці глікогенфосфорилаза активується внаслідок фосфорилювання. Фосфорильовану її форму називають фосфорилазою a, а не фосфорильовану (і, відповідно, неактивну) — фосфорилазою b. Перетворення b форми в a забезпечує кіназа фосфорилази. Вона у свою чергу також є регуляторним ферментом і може активуватись під впливом фосфорилювання або/і підвищеної концентрації кальцію в цитоплазмі. Фосфорилює кіназу фосфорилази протеїнкіназа А, яка функціонує тільки за наявності певного рівня цАМФ. Збільшення концентрації вторинного посередника цАМФ у клітині відбувається під дією гормонів, глюкагону та адреналіну (коли він зв'язується із β-адренергічним рецептором)[13][14].
Збільшення концентрації кальцію, який також відіграє роль в активації кінази фосфорилази, у гепатоцитах також відбувається під впливом гормону адеренліну, проте в тому випадку, коли він діє через α-адернорецептор. У скелетних м'язах таких рецепторів немає, проте рівень кальцію зростає безпосередньо у відповідь на нервовий імпульс, що стимулює скорочення[15].
Зворотне перетворення глікогенфосфорилази a у глікогенфосфорилазу b, а також і дефосфорилювання та інактивацію кінази фосфорилази, забезпечує фосфопротеїнфосфатаза 1. Активність цього ферменту пригнічується у відповідь на дію глюкагону та адреналіну. Після припинення гормонального сигналу глікогеноліз швидко «вимикається»: конститутивно активна фосфодіестераза гідролізує цАМФ, протеїнкіназа А інактивуються, кіназа фосфорилази та глікогенфосфорилаза дефосфорилюється фосфопротеїнфосфатазою 1[16].
Пригнічення глікогенолізу
Коли м'язова тканина достатньо забезпечена енергією та енергетичними сусбтратами, відбувається миттєве сповільнення глікогенолізу. АТФ та глюкозо-6-фосфат діють як алостеричні інгібітори глікогенфосфосфорилази[11].
Глікогенфосфорилаза печінки, а саме її активна a форма, чутлива до концентрації глюкози. Після вживання їжі її рівень в крові зростає і вона переноситься транспортером GLUT2 у гепатоцити, де діє як алостеричний інгібітор глікогенфосфорилази. Активна фосфорилаза a пригнічувала фосфопротеїнфосфатазу 1, а після її інактивації глюкозою блокування знімається і фосфопротеїнфосфатаза 1 дефосфорилює фосфорилазу a та переводить її в неактивну b форму[17].
Глікогеноліз у м'язах та печінці також пригнічується гормоном інсуліном, що виділяється підшлунковою залозою у відповідь на високі концентрації глюкози. Інсулін не тільки блокує розщеплення глікогену а одночасно стимулює його біосинтез[18].
Примітки
- Berg et al, 2007, с. 594.
- Nelson et al, 2008, с. 595.
- Voet et al, 2011, с. 640.
- Nelson et al, 2008, с. 596.
- Voet et al, 2011, с. 639.
- Berg et al, 2007, с. 595.
- Voet et al, 2011, с. 646.
- Berg et al, 2007, с. 596.
- Voet et al, 2011, с. 642.
- Berg et al, 2007, с. 598.
- Berg et al, 2007, с. 599.
- Voet et al, 2011, с. 648.
- Nelson et al, 2008, с. 603.
- Berg et al, 2007, с. 600—603.
- Berg et al, 2007, с. 603.
- Berg et al, 2007, с. 603—604.
- Berg et al, 2007, с. 610.
- Nelson et al, 2008, с. 608.
Джерела
- Berg JM, Tymoczko JL, Stryer L (2007). Biochemistry (вид. 6th). W.H. Freeman and Company. ISBN 0-7167-8724-5.
- Nelson D.L., Cox M.M. (2008). Lehninger Principles of Biochemistry (вид. 5th). W. H. Freeman. ISBN 978-0-7167-7108-1.
- Voet D., Voet J.G. (2011). Biochemistry (вид. 4th). Wiley. с. 487—496. ISBN 978-0470-57095-1.
