Інсулін
Інсулін (від лат. insula — острів) — гормон пептидної природи, що утворюється у бета-клітинах острівців Лангерганса підшлункової залози. Впливає на багато аспектів обміну речовин практично у всіх тканинах. Основна дія інсуліну полягає в зниженні концентрації глюкози в крові.
| Інсулін | |
|---|---|
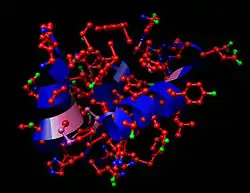 Структура молекули інсуліну | |
| Ідентифікатори | |
| Символ | INS |
| DrugBank | |
| Entrez | |
| HUGO | |
| OMIM | |
| PDB | 1AIY |
| UniProt | |
| Інша інформація | |
| Локус | Хр. 11 |
Інсулін збільшує проникність плазматичних мембран для глюкози, активує ключові ферменти гліколізу, стимулює перетворення в печінці і м'язах глюкози на глікоген, підсилює синтез жирів і білків. Крім того, інсулін пригнічує активність ферментів, що розщеплюють глікоген і жири. Тобто, крім анаболічної дії, інсулін володіє також і антикатаболічним ефектом.
Порушення секреції інсуліну унаслідок деструкції бета-клітин — абсолютна недостатність інсуліну — є ключовою ланкою патогенезу цукрового діабету 1-го типу. Порушення дії інсуліну на тканини — відносна інсулінова недостатність — має важливе місце в розвитку цукрового діабету 2-го типу.
Будова молекули інсуліну
Інсулін — це невеликий білок молекулярною масою 5,8 кДа. Він складається із двох поліпептидних ланцюгів: А (21 амінокислота) та B (30 амінокислот). Молекула інсуліну містить три дисульфідні зв'язки: два із них сполучають між собою A та B-ланцюги, а третій розташований всередині A-ланцюга. Бичачий інсулін має таку первинну структуру[1]:
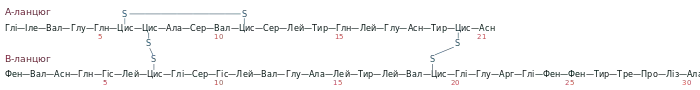
Структура інсуліну майже не змінилась в еволюції вищих хребетних, зокрема інваріабельними є положення дисульфідних зв'язків, аміно- та карбокситермінальні ділянки A-ланцюга, та гідрофобні амінокислоти близько C-кінця B-ланцюга[2]. Людський інсулін відрізняється від бичачого двома амінокислотними замінами в A-ланцюгу: у 8-му положенні треонін замість аланіну, а у 10-му ізолейцин замість валіну. Свинячий гормон ще ближчий до людського, він відрізняється всього однією амінокислотою: аланіном у 30-му положенні B-ланцюга замість треоніну[3].
У розведеному розчині молекули інсуліну існують в мономерному стані, кожна така молекула складається із гідрофобної серцевини та переважно гідрофільної поверхні, за винятком двох неполярних ділянок. Ці ділянки беруть участь в утворенні димерів та гексамерів. У концентрованих розчинах, наприклад в препаратах для ін'єкції, та кристалах, як всередині секреторних везикул β-клітин, шість мономерів інсуліну разом із двома атомами Цинку утворюють гексамер. Таким чином після підшкірного введення інсуліну він всмоктується у кров повільно, через те, що для дисоціації гексамерів необхідний додатковий час[4].
Утворення і секреція
Синтез інсуліну в клітині
 |
 |
 | |
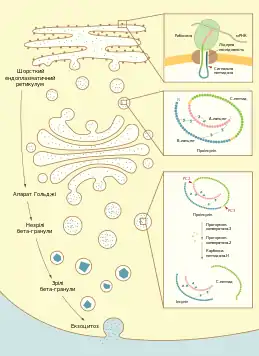
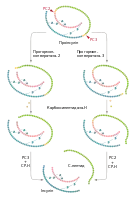
| Біосинтез інсуліну. Зліва: загальна схема біосинтезу. Справа зверху: схема обмеженого протеолізу проіснуліну з утворенням інсуліну та C-пептиду. Справа знизу: бета-клітина пацюка з великою кількістю гранул заповнених інсуліном (ТЕМ) | |
Інсулін синтезується у β-клітинах острівців Лангерганса підшлункової залози. Ген попередника інсуліну — препроінсуліну — в людини локалізується в короткому плечі 11 хромосоми. Він містить 3 екзони та 2 інтрони[5]. В інших тварин, наприклад мишей, пацюків та трьох видів риб, наявні два гени інсуліну[6].
Препроінсулін людини складається із 110 амінокислот: 24 з них становлять гідрофобну N-кінцеву лідерну послідовність (сигнальний пептид), за нею розташований B-ланцюг, далі — послідовність Арг-Арг, з'єднувальний C-пептид (від англ. connecting peptide — з'єднувальний пептид), послідовність Ліз-Арг, та А-ланцюг на C-кінці[2]. Лідерна послідовність необхідна для котрансляційного транспорту препроінсуліну в порожнину шорсткого ендоплазматичного ретикулуму[5][7]. Після проходження через мембрану лідерна послідовність відщеплюється спеціальною сигнальною пептидазою[8] і швидко деградує. Утворений після цього проінсулін складається із 86 амінокислотних залишків і не має гормональної активності. У ендоплазматичному ретикулумі відбувається його згортання та формування всередині молекули трьох дисульфідних зв'язків[9].
Після утворення правильної просторової структури проінсулін у транспортних везикулах переноситься до цис-сторони комплексу Гольджі. В ході руху прогормону від цис- до транс-Гольджі відбувається його відсортовування в компартмент секреторних гранул. Тут, у незрілих гранулах, проінсулін підлягає подальшій модифікації, а саме обмеженому протеолізу, що починається із дії двох прогормонконвертаз (PC2 і PC3). Ці ферменти діють специфічно на карбоксикінцевій стороні послідовності з двох позитивно заряджених амінокислот. В молекулі проінсуліну є два таких сайти: Арг31-Арг32 (місце дії PC2) та між Ліз64-Арг65 (місце дії PC3), де і відбувається розрив пептидних зв'язків. Відразу ж після прогормонконвертаз ферментативну активність проявляє карбоксипептидаза-H, яка відщеплює основні амінокислоти від утворених кінців. Кінцевими продуктами протеолізу є молекула інсуліну та C-пептид довжиною 31 амінокислота[10][11]. Порівняно із A- та B-ланцюгами інсуліну C-пептид є значно більш варіабельним у хребетних тварин, його довжина коливається від 28 (у корів) до 38 у представників родини Вудильникові[2].
Зрілі секреторні везикули β-клітин містять кристалічний інсулін у формі гексамерів з атомами цинку та еквімолярну кількість C-пептиду. Вони становлять пул гормону, готовий до екзоцитозу у відповідь на стимул. Час півжиття β-гранул становить кілька днів, і якщо вони не секретують свій вміст, то підлягають деградації шляхом злиття із лізосомами. При підвищеній потребі організму в інсуліні деградація відбувається повільніше[12].
Регуляція синтезу інсуліну відбувається на кількох рівнях, зокрема на рівні транскрипції, сплайсингу пре-мРНК, деградації мРНК[13], трансляції та посттрансляційної модифікації[14]. Найсильнішим стимулятором цих процесів є глюкоза, проте біосинтез проінсуліну може активуватись також іншими цукрами, амінокислотами, зокрема лейцином, проміжними продуктами гліколізу, кетоновими тілами, гормоном росту, глюкагоном та деякими іншими факторами[15].
Секреція інсуліну
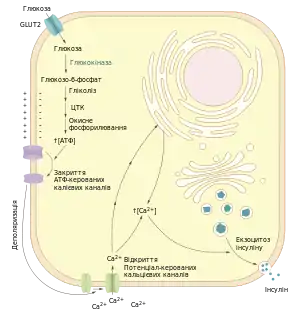
Бета-клітини підшлункової залози, як типові ендокринні клітини, секретують більшість (95%) свого основного продукту — інсуліну — регульованим шляхом[16][17]. Найважливішим активатором цього шляху є глюкоза. У мембранах бета-клітин постійно наявні переносники глюкози GLUT2, через які вона може вільно дифундувати. Завдяки цьому збільшення концентрації глюкози в крові призводить до аналогічного підвищення її рівня і в бета-клітинах. Тут вона відразу ж стає субстратом гексокіназної реакції, продуктом якої є глюкозо-6-фосфат. В інсулін-синтезуючих клітинах підшлункової залози експресується один із ізоферментів гексокінази — гексокіназа IV або глюкокіназа[18][17], для неї характерна низька спорідненість до субстрату: константа Міхаеліса становить 10 мМ, що перевищує нормальний вміст глюкози в крові (4-5 мМ)[19]. Завдяки цьому глюкокіназа може працювати «глюкозним сенсором», активуючись тільки в умовах гіперглікемії[17].
Глюкозо-6-фосфат вступає в реакції гліколізу, продукти якого далі окиснюються у мітохондріях, внаслідок чого в клітині утворюється велика кількість АТФ. Підвищення концентрації АТФ призводить до закриття АТФ-керованих калієвих каналів (англ. ATP-gated K+ channels, KATP) у плазмалемі. Внаслідок зменшення відтоку калію із клітини мембрана деполяризується, а це веде до відкриття потенціал-керованих кальцієвих каналів і притоку кальцію в клітину. Початкове збільшення концентрації іонів Ca2+ у цитозолі веде до подальшого їх вивільнення із ендоплазматичного ретикулуму. Кальцій викликає злиття клатрин-облямованих бета-гранул із плазмалемою і вивільнення їх вмісту в міжклітинний простір[18][17], звідки інсулін потрапляє в кров через фенестровані стінки капілярів[5].
На активність АТФ-керованих калієвих каналів окрім власне АТФ можуть впливати також інші речовини. Ці трансмембранні білки складаються із восьми субодиниць: чотирьох ідентичних Kir6.2 та чотирьох ідентичних SUR1. Перші формують гідрофільний тунель і відповідають за чутливість до АТФ, а другі є рецепторами до сульфанілсечовин (англ. sulphonylurea receptor) і можуть інактивувати канал після зв'язування зі своїм лігандом. Таким чином сульфанілсечовини активують синтез інсуліну, завдяки чому використовуються як пероральні цукрознижувальні препарати при цукровому діабеті[20][17].
Окрім регульованого існує так званий «конститутивний шлях» секреції інсуліну бета-клітинами, він працює за певних розладів, таких як інсулінома та цукровий діабет другого типу. В цьому випадку велика кількість незрілого гормону (проінсуліну або проміжних «розщеплених» форм) виділяється прямо із везикул, що утворюються в ендоплазматичному ретикулумі[17].
Регуляція секреції інсуліну
Острівці Лангерганса густо іннервовані автономними та пептидергічними нервовими волокнами. Холінергічні закінчення блукаючного нерва, що є частиною парасимпатичної нервової системи стимулюють секрецію інслуіну, в той час як адренергічні закінчення симпатичної нервової системи пригнічують цей процес. Інші нерви виділяють вазоактивний інтестинальний пептид, що стимулює секрецію всіх гормонів підшлункової залози, та нейропептид Y, що блокує виділення інсуліну[21].
Власні гормони підшлункової залози також мають вплив на секрецію інсуліну: глюкагон стимулює її, а соматостатин — пригнічує. Окрім того інсулін діє автокринно активуючи транскрипцію власного гену і гену глюкокінази[21].
Під час прийому їжі секреція інсуліну збільшується не тільки під впливом глюкози або вуглеводів, а також амінокислот, особливо лейцину і аргініну, деяких гормонів травної системи системи: холецистокініну, глюкозозалежного інсулінотропного пептиду, а також таких гормонів, як глюкагон, адренокортикотропний гормон, естроген та інші. Також секрецію інсуліну підсилює підвищення рівня калію або кальцію, вільних жирних кислот в плазмі крові.
Інкертиновий ефект
Інкретиновий ефект — це феномен, що полягає виділенні значно більшої кількістю інсуліну, у відповідь на пероральне вживання глюкози у порівнянні із її внутрішньовенним введенням. За це явище відповідають гормони травного тракту, що секретуються під час вживання їжі і посилюють глюкозо-стимульоване вивільнення інсуліну. До інкретинових гормонів належать зокрема глюкагоноподібний пептид-1 та шлунковий інгібуючий поліпептид, перший із них секретуються L-, а другий — K-клітинами верхньої частини порожньої кишки[22].
Фізіологічна дія інсуліну
|
Метаболічний ефект |
Молекула-мішень |
|
↑ Захоплення глюкози (м'язи та жирова тканина) |
↑ Транспортер глюкози GLUT4 |
|
↑ Захоплення глюкози (печінка) |
↑ Глюкокіназа (підвищена експресія) |
|
↑ Синтез глікогену (печінка та м'язи) |
|
|
↓ Розщеплення глікогену (печінка та м'язи) |
|
|
↑ Гліколіз, продукування ацетил-КоА (печінка та м'язи) |
↑ Фосфофруктокіназа-1 (через ↑ФФК-2) |
|
↑ Синтез жирних кислот (печінка) |
↑ Ацетил-коА-карбоксилаза |
|
↑ Синтез триацилгілцеролів (жирова тканина) |
Основна фізіологічна дія інсуліну полягає у зниженні вмісту глюкози в крові, проте вона не обмежується цим, гормон також впливає на метаболізм білків та ліпідів. Одним із визначальних ефектів інсуліну є те, що він стимулює посилення засвоєння глюкози м'язами та жировою тканиною, проте не впливає на цей процес у печінці, нирках та мозку, клітини яких можуть транспортувати глюкозу навіть за відсутності гормональної стимуляції. Також інсулін блокує ті метаболічні шляхи, кінцевим продуктом яких є глюкоза, зокрема глюконеогенез та розщеплення глікогену, і стимулює ті, в яких вона використовується. Перший пріоритет при цьому належить задоволенню енергетичних потреб, зокрема протіканню гліколізу, кінцевим продуктом якого є піруват, та подальшого окиснення пірувату до ацетил-КоА, який може бути використаний у циклі трикарбонових кислот. Залишок глюкози використовується на поповнення запасів глікогену у печінці та м'язах. У печінці інсулін також стимулює синтез жирних кислот із ацетил-коА, необхідний для цього НАДФH виробляється у пентозофосфатному шляху. Далі жирні кислоти у формі тригліцеридів транспортуються до жирової тканини[24][25]. Інсулін також впливає на метаболізм опосередковано через головний мозок. Він впливає на ядра гіпоталамуса таким чином, що пригнічує споживання їжі та посилює термогенез[26].
У м'язовій тканині інсулін стимулює захоплення амінокислот і синтез білків[24]. Залишок амінокислот перетворюється у печінці до пірувату та ацетил-КоА і використовується для синтезу жирів[25].
Поглинання кальцію клітинами також активується під впливом інсуліну. Тому його препарати разом із глюкозою використовують для тимчасового зниження гіперкальціємії у пацієнтів із нирковою недостатністю. Точний молекулярний механізм такої дії інсуліну не з'ясований, проте відомо, що він може активувати Na+/K+-АТФазу[27].
До довготривалих ефектів інсуліну на організм належить прискорення росту, що відбувається завдяки його загальному анаболічному та білок-заощаджувальному впливу. Тому у дітей із цукровим діабетом першого типу спостерігається затримка росту. Інсулін може стимулювати ріст незрілих гіпофізектомічних щурів, майже з такою ж інтенсивністю як і гормон росту за умови, якщо вони вживають велику кількість вуглеводів[28]. Також відомо, що в культурі клітин інсулін пришвидшує клітинний поділ, подібно до пептидних факторів росту, таких фактор росту епідермісу, фактор росту фібробластів та тромбоцитарний фактор росту, і, окрім того, може посилювати їхній біологічний вплив[29].
Пов'язані захворювання
Проте в контексті проблеми, що розглядається, слід наголосити на особливій ролі інсуліну в регуляції енергетичного метаболізму в цілому, включаючи обмін не лише вуглеводів, а й жирів. Це стосується механізмів координованого збереження й утилізації паливних молекул у жировій тканині, печінці та скелетних м'язах. Після перетравлення їжі в організм надходить велика кількість вуглеводів, проте їх концентрація у периферичній крові та міжклітинному просторі завдяки активації екскреції інсуліну не досягає критичних величин. Цей гормон стимулює надходження глюкози в інсулінозалежні органи і тканини й одночасно пригнічує процеси утворення ендогенної глюкози завдяки супресії глюконеогенезу та глікогенолізу. Паралельно інсулін стимулює і синтез глікогену. Подібну дію інсулін також проявляє щодо обміну жирів. Це маніфестується стимуляцією процесів накопичення жиру в жировій тканині й пригніченням мобілізації жиру з депо внаслідок активації інсуліном ліпопротеїнової ліпази, що сприяє очищенню крові від тригліцеридів та інгібіції активності гормоночутливої ліпази. Одночасно інсулін стимулює надходження в ліпоцити глюкози та збуджує синтез внутрішньоклітинних тригліцеридів, тобто активує ліпогенез. У цьому плані жирова тканина виконує своєрідну буферну функцію, забезпечуючи нормалізацію концентрації жирів у плазмі, особливо в постпрандіальний період. У стані спокою або короткочасного голоду концентрація інсуліну в крові знижується, підвищується рівень контрінсулярних гормонів та стимулюється функція симпатичної нервової системи, що призводить до мобілізації печінкової глюкози та активації ліполізу з вивільненням із адипоцитів у кровообіг неетерифікованих ВЖК. За цих умов глюкоза переважно утилізується такими інсулінонезалежними тканинами, як нервова система та еритроцити, тоді як скелетні м'язи одержують енергію і за рахунок окислення жирних кислот. За подовженого стану голоду в печінці додатково відбувається перетворення жирних кислот у кетонові тіла, а ацетил-КоА — у глюкозу. Подібні зміни мають місце і при фізичному навантаженні організму, але з посиленням надходження глюкози в м'язи. Отже, накопичення і витрачання жирів є динамічним процесом, що протікає по-різному залежно від стану організму та його потреб в енергії. Слід також зауважити, що інсулін — не єдиний гормон, який регулює енергетичний обмін. Існує ціла низка контрінсулярних гормонів (глюкагон, адреналін, гормон росту і глюкокортикоїди), активність яких спрямовано на збільшення в циркуляції концентрації глюкози. Додатковим антагоністом інсуліну виступає симпатична нервова система, стимуляція якої призводить до вивільнення ВЖК із жирових клітин. Наявність такого складного регуляторного механізму передбачає можливість ушкодження або зниження чутливості тканин до регуляторних чинників процесів накопичення та утилізації енергетичних ресурсів на різних рівнях.
Використання інсуліну як ліків
При цукровому діабеті.
Відкриття і дослідження
Відкриття ендокринної функції підшлункової залози

У 1869 році в Берліні 22-річний студент-медик Пауль Лангерганс досліджуючи за допомогою нового мікроскопу будову підшлункової залози, звернув увагу на раніше не відомі групи клітини рівномірно розподілені по її тканині. Він не робив жодних припущень щодо їх призначення. Тільки 1893 року Едуард Лагес з'ясував, що вони відповідають за ендокринну функцію підшлункової залози і назвав їх «острівцями Лангерганса» в честь першовідкривача[30].
У 1889 році Оскар Мінковський та Йозеф фон Мерінг досліджували функції підшлункової залози. Вони провели панкреатомію здоровому собаці, через декілька днів після цього в тварини почали спостерігатись симптоми діабету, зокрема сильна спрага, посилене виділення сечі із високим вмістом цукру, надмірне споживання їжі та втрата ваги. Через деякий час дослідникам вдалось «вилікувати» собаку шляхом трансплантації тканини підшлункової залози під шкіру[31][32].
У 1901 році був зроблений наступний важливий крок, Юджин Опі (Eugene Opie) чітко показав, що «цукровий діабет обумовлений руйнуванням острівців підшлункової залози, і виникає тільки коли ці тільця частково або повністю зруйновані»[32]. Зв'язок між цукровим діабетом і підшлунковою залозою був відомий і раніше, але до цього не було ясно, що діабет пов'язаний саме з острівцями Лангерганса. Це, а також багато інших досліджень патологій підшлункової залози, привело таких вчених як Жан де Меєр (1909)[33][31] та Едвард Шарпі-Шафер (1916)[32] до висновку про те, що острівці Лангерганса повинні виробляти речовину із цукрознижувальним ефектом. Мейер назвав її інсуліном від лат. insula — острів[31].
Перші спроби виділення інсуліну
У наступні два десятиліття було зроблено декілька спроб виділити острівцевий секрет як потенційні ліки. У 1907 році Георг Зюльцер (Georg Ludwig Zülzer) досяг деякого успіху в зниженні рівня глюкози в крові піддослідних собак панкреатичним екстрактом і навіть зміг врятувати одного пацієнта у діабетичній комі. Проте його препарат мав сильні побічні ефекти, ймовірну, через погане очищення, через що від нього довелось відмовитись[31].
Ернест Скотт між 1911 і 1912 роками в Чиказькому університеті використовував водний екстракт підшлункової залози і відзначав «деяке зменшення глюкозурії», але він не зміг переконати свого керівника у важливості цих досліджень, і незабаром експерименти були припинені. Такий же ефект демонстрував Ізраель Кляйнер в Рокфеллерівському університеті в 1919 році, але його робота була перервана початком Першої світової війни, і він не зміг її завершити. Схожу роботу після дослідів у Франції в 1921 році опублікував професор фізіології Румунської школи медицини Ніколас Паулеско, і багато хто, особливо в Румунії, вважають саме його першовідкривачем інсуліну.
Робота Бантинга та Беста



Більшість екстрактів підшлункової залози, виготовлених різними дослідниками у період до 1921-го року, мали одну й ту ж проблему: вони містили багато домішок, зокрема продуктів екзокринної частини залози, і викликали абсцеси у пацієнтів. Вперше виділити та очистити до рівня придатного до використання для терапії людей вдалось групі вчених Торонтського університету у 1921 році[34].
Фредерик Бантінг після завершення Першої світової війни працював хірургом-ортопедом та читав лекції в Університеті Західного Онтаріо. Однією із тем цих лекцій був метаболізм вуглеводів. В ході ознайомлення із предметом, Бантинг прочитав працю доктора Мозеса Баррона, в якій той описував відмирання екзокринної частини підшлункової залози у випадках, коли її протока була перекрита камінцями[35]. Це навело його на думку про новий метод виділення ендокринного секрету підшлункової залози, у своїх нотатках Бантінг записав[36]:
Перев'язати панкреатичну протоку собаки. Підтримувати життєдіяльність тварини поки ацинуси не дегенерують і залишаться тільки острівці. Спробувати виділити внутрішній секрет і подіяти на глікозурію.
Зі своєю ідеєю Бантинг звернувся до Джона Маклеода — професора Університету Торонто, міжнародно відомого дослідника метаболізму вуглеводів. Маклеод знав про труднощі, з якими зіткнулись попередні дослідники у спробах виділити лікувальний панкреатичний екстракт, проте він вважав, що навіть негативний результат роботи Бантинга буде корисним, і тому погодився надати йому місце в лабораторії, собак для дослідів та одного помічника[35][37]. На роль помічника претендували два студенти фізіологи: Чарльз Бест та Кларк Ноубл. Щоб вирішити, хто саме з них допомагатиме Бантингу, вони кинули монетку. Хоча поширеною є думка про те, що Бест виграв, його знайомий Роберт Волп наполягав на тому, що він програв, оскільки жоден із студентів не хотів працювати із меланхолінйим дратівливим Бантингом[38].
Влітку 1921 Бантінгу та Бест розпочали свої експерименти і після невеликої затримки отримали атрофовані підшлункові залози. Екстракт отримували таким чином: різали тканину на шматки, розтирали її у ступці та фільтрували розчин, після чого вводили діабетичним собакам[36]. Хоча їм вдалось добитись зниження рівня глюкози в крові тварин, Бантинг та Бест зіткнулись із тією ж проблемою, що й їхні попередники: на місці ін'єкції розвивався стерильний абсцес і загальна інтоксикація. Впродовж кінця літа та осені 1921 року вони виявили, що екстракт із атрофованої підшлункової залози собак не мав ніякої переваги над екстрактом із фетальних залоз телят з бійні. І тому стали надалі використовувати цю тканину, яку було значно легше отримати[38].
Наприкінці 1921 року Маклеод запропонував біохіміку Джеймсу Колліпу приєднатись до групи Бантинга та Беста і попрацювати над новими методами очищення екстракту. Колліп погодився, і згодом також показав, що панкреатичний екстракт стимулює відкладання глікогену в печінці, зменшує кетоацидоз в діабетичних тварин, та знижує рівень глюкози в крові у здорових[39]. Після цього для тестування перепарату він використовував нормальних кроликів, а не панкреатомованих собак. У листопаді 1921 року Бантинг та Маклеод відвідали збори Американського фізіологічного товариства, де представили отримані результати[36].
11 січня 1922 року Бантинг та Бест вперше випробували один із активних екстрактів телячої підшлункової залози, який вони назвали «айлетином» від англ. isle — острів, на пацієнті — 14-річному Леонарді Томпсоні. Йому ввели по 7,5 мл перепарату у кожен сідничний м'яз і отримали очікуваний результат: рівень глюкози в крові знизився, проте розвинувся абсцес та загальне отруєння. Через кілька тижнів після цієї невдачі, Колліп повідомив Бантингу, що йому врешті вдалось отримати нетоксичний екстракт, але не захотів розповісти деталі процесу (ймовірно, сподіваючись на майбутній патент), що мало не призвело до бійки між дослідниками[40].
Новому екстракту Колліпа Маклеод дав назву «інсулін» (ймовірно, не підозрюючи, що таку ж назву використовував Меєр 1909 року). Його випробували 23 січня на тому ж пацієнті. Цього разу лікування було успішним: рівень глюкози в крові Томпсона впав від 520 до 120 мг/дл, і не спостерігалось ніяких побічних ефектів[36]. Але згодом з'ясувалось, що Колліп забув протокол приготування інсуліну. Впродовж наступних кількох тижнів із допомогою Елі Ліллі він намагався знову віднайти метод, що йому, зрештою, вдалось зробити[40].
Серед перших пацієнтів, яких лікували Бантинг та Бест була Елізабет Х'юз дочка державного секретаря Чарльза Х'юза. Вона описала зміну свого стану здоров'я після ін'єкції інсуліну як «несказанно чудове». Відомі діабетологи того часу Елліот Джослін та Фредерік Аллен також були вражені силою нового препарату. Описуючи свої враження Джослін порівняв дію інсуліну зі сценою з Біблії Єз. 37:1-10[41][42]:
Була надо мною Господня рука, і Дух Господній випровадив мене, і спинив мене серед долини, а вона повна кісток! ... вони стали дуже сухі! І сказав Він мені: Сину людський, чи оживуть оці кості? А я відказав: Господи Боже, Ти знаєш! І сказав Він мені: Пророкуй про ці кості, та й скажеш до них: Сухі кості, послухайте слова Господнього! ...І пророкував я, як наказано. І знявся шум, коли я пророкував, і ось гуркіт, а кості зближалися, кістка до кістки своєї. І побачив я, аж ось на них жили, і виросло тіло, і була натягнена на них шкіра зверху, та духа не було в них. І сказав Він мені: Пророкуй до духа, пророкуй, сину людський, та й скажеш до духа: Так говорить Господь Бог: Прилинь, духу, з чотирьох вітрів, і дихни на цих забитих, і нехай оживуть! І я пророкував, як Він наказав був мені, і ввійшов у них дух, і вони ожили, і поставали на ноги свої, військо дуже-дуже велике!...
За революційне відкриття інсуліну Маклеод і Бантінг в 1923 році були удостоєні Нобелівської премії з фізіології і медицини. Бантінг спершу був сильно обурений, що його помічник Бест не був представлений до нагороди разом з ним, і спочатку навіть демонстративно відмовився від нагороди, але потім все ж таки погодився прийняти премію, і свою частину урочисто розділив з Бестом. Так само поступив і Маклеод, поділивши свою премію з Колліпом. Патент на виробництво телячого інсуліну отримали Бест та Колліп, як немедичні члени групи, оскільки участь медиків у комерційних справах вважалась неетичною. Вони передали патент Торонтському університету, а він надав ліцензії багатьом медичним фірмам по всьому світу. Зокрема у США право на виробництво інсуліну отримала фармацевтична фірма Елі Ліллі, у Європі найбільшим виробником стала корпорація заснована в Данії Августом Крогом[43].
Дослідження структури інсуліну
Інсулін був першою білковою молекулою, для якої було повністю встановлено амінокислотну послідовність, тобто первинну структуру. Цю роботу здійснив 1953 року британський молекулярний біолог Фредерік Сенгер, за що був удостоєний Нобелівської премії з хімії 1958 року. А через майже 40 років Дороті Кроуфут Ходжкін за допомогою методу рентгенівської дифракції визначила просторову будову (третинну структуру) молекули інсуліну. Її роботи також відзначені Нобелівською премією[44].
Штучне виробництво інсуліну
Починаючи із 1980-их років людський інсулін отримують генноінженерними методами за допомогою клітин кишкової палички або пивних дріжджів[44]. В Україні виробництво інсуліну наявне на ПрАТ "Індар".
Див. також
Примітки
- Voet et al, 2011, с. 165.
- Rhodes, 2004, с. 28.
- Bilous et al, 2010, с. 7.
- Bilous et al, 2010, с. 25—26.
- Ґаноґ, 2002, с. 307.
- Insulin у базі даних OMIM — Переглянуто 10 липня 2012 (англ.)
- Березов et al, 1998, с. 268.
- Voet et al, 2011, с. 1404.
- Rhodes, 2004, с. 35.
- Bilous et al, 2010, с. 26.
- Rhodes, 2004, с. 38.
- Rhodes, 2004, с. 40.
- German, 2004, с. 21.
- Rhodes, 2004, с. 31—33.
- Rhodes, 2004, с. 32.
- Rhodes, 2004, с. 41.
- Bilous et al, 2010, с. 28.
- Nelson et al, 2008, с. 924.
- Nelson et al, 2008, с. 584—585.
- Nelson et al, 2008, с. 925.
- Bilous, 2010, с. 23.
- Bilous, 2010, с. 28.
- Nelson et al, 2008, с. 923.
- Marieb, 2006
- Nelson et al, 2008, с. 922—923.
- Nelson et al, 2008, с. 934.
- Ґаноґ, 2002, с. 309.
- Ґаноґ, 2002, с. 311.
- Губський, 2007, с. 425—426.
- Bilous et al, 2010, с. 6.
- Zajac et al, 2010, с. 5.
- Eknoyan G, Nagy J. (2005). A history of diabetes mellitus or how a disease of the kidneys evolved into a kidney disease. Adv Chronic Kidney Dis 12: 223–9. PMID 15822058. doi:10.1053/j.ackd.2005.01.002.
- Ali H., Anwar M, Ahmad T., Chand N. (2006). Diabetes Mellitus from Antiquity to Present Scenario and Contribution of Greco-Arab Physicians. JISHIM 5: 46–50.
- Strakosch, 2000, с. 7.
- Strakosch, 2000, с. 8.
- Zajac et al, 2010, с. 6.
- Bliss, 1997, с. 93.
- Strakosch, 2000, с. 9.
- Bliss, 1997, с. 95.
- Strakosch, 2000, с. 11.
- Bliss, 1997, с. 98.
- Strakosch, 2000, с. 12.
- Strakosch, 2000, с. 15.
- Zajac et al, 2010, с. 8.
Джерела
- Bilous, Rudy W (2010). Handbook of diabetes (вид. 4th). Wiley-Blackwell. ISBN 978-1-4051-8409-0.
- Zajac J, Shrestha A, Patel P, Poretsky L (2010). The Main Events in the History of Diabetes Mellitus. У Poretsky L. Principles of Diabetes Mellitus (вид. 2nd). New York: Springer. ISBN 978-0-387-09840-1. doi:10.1007/978-0-387-09841-8.
- Strakosch C (2000). The discovery of Insulin. Ramsay Health Care.
- Bliss M (1997). The Discovery of Insulin: The Inside Story. У Higby G., Stroud E. The Inside Story of Medicines: A Symposium. American Institute of History of Pharmacy. ISBN 0-931292-32-8.
- Rhodes CJ (2004). Processing of the Insulin Molecule. У LeRoith D, Taylor SI, Olefsky JM. Diabetes mellitus: a fundamental and clinical text (вид. 3rd). American Institute of History of Pharmacy. ISBN 0-7817-4097-5. Архів оригіналу за 4 листопада 2013. Процитовано 11 липня 2012.
- German M (2004). Insulin Gene Regulation. У LeRoith D, Taylor SI, Olefsky JM. Diabetes mellitus: a fundamental and clinical text (вид. 3rd). American Institute of History of Pharmacy. ISBN 0-7817-4097-5. Архів оригіналу за 21 лютого 2009. Процитовано 21 липня 2012.
- Voet D., Voet J.G. (2011). Biochemistry (вид. 4th). Wiley. с. 487—496. ISBN 978-0470-57095-1.
- Berg JM, Tymoczko JL, Stryer L (2007). Biochemistry (вид. 6th). W.H. Freeman and Company. ISBN 0-7167-8724-5.
- Nelson D.L., Cox M.M. (2008). Lehninger Principles of Biochemistry (вид. 5th). W. H. Freeman. ISBN 978-0-7167-7108-1.
- Marieb EN, Hoehn K (2006). Human Anatomy & Physiology (вид. 7th). Benjamin Cummings. ISBN 978-0805359091.
- Березов Т. Т., Коровкин Б. Ф. (1998). Биологическая химия: Учебник. (вид. 3). Москва: Медицина. с. 704. ISBN 5-225-02709-1.
- Губський Ю.І. (2007). Біологічна хімія. Київ-Вінниця: Нова книга. с. 656. ISBN 978-966-382-017-0.
- Ґаноґ В.Ф. (2002). Фізіологія людини [Гжегоцький М., Шевчук В., Заячківська О.]. Львів: БаК. с. 784. ISBN 966-7065-38-3.
Посилання
- ІНСУЛІН //Фармацевтична енциклопедія
- David Goodsell Insulin Molecule of the Month Feb 2001 — Переглянуто 10 липня 2012 (англ.)