Перемикання класу імуноглобулінів
Перемикання (світчінг) класу імуноглобуліну (антитіла), також відоме як перемикання ізотипу, ізотипна комутація або рекомбінація з перемиканням класів (CSR), є біологічним механізмом, який змінює вироблення В-клітинами імуноглобуліну з одного типу на інший, наприклад з ізотипу IgM на ізотип IgG.[1] Під час цього процесу частина важкого ланцюга антитіла з постійною (константною) ділянкою змінюється, але варіабельна область важкого ланцюга залишається незмінною (терміни варіабельна і постійна відносяться до змін або їх відсутності між антитілами, які націлені на різні епітопи). Оскільки варіабельна ділянка не змінюється, перемикання класів не впливає на специфічність антигену. Натомість антитіло зберігає спорідненість до тих самих антигенів, але може взаємодіяти з різними ефекторними молекулами.
Механізм
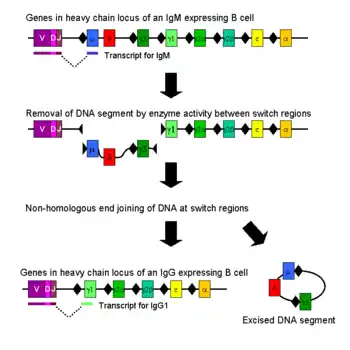
Перемикання класів відбувається після активації зрілої В-клітини через її мембранозв’язане антитіло (або В-клітинний рецептор) для створення різних класів антитіл, з тими ж варіабельними доменами, що й вихідне антитіло, що генерується в незрілій В-клітині під час процесу рекомбінації V(D)J. Змінюються константні домени у важких ланцюгах.[2]
Наївні зрілі В-клітини виробляють як IgM, так і IgD, які є першими двома сегментами важкого ланцюга в локусі імуноглобуліну. Після активації антигеном ці В-клітини проліферують. Якщо ці активовані В-клітини стикаються зі специфічними сигнальними молекулами через свої CD40 і цитокінові рецептори (обидва модулюються Т-хелперами ), вони піддаються перемиканню класу антитіл для вироблення антитіл IgG, IgA або IgE. Під час зміни класу антитіла константна ділянка важкого ланцюга імуноглобуліну змінюється, але варіабельні ділянки - ні, а отже, антигенна специфічність залишаються незмінними. Це дозволяє різним дочірнім клітинам з однієї активованої В-клітини виробляти антитіла різних ізотипів або підтипів (напр. IgG1, IgG2 тощо).[3]
У людей виділяються таки порядок екзонів важких ланцюгів:
Перемикання класів відбувається за допомогою механізму, який називається зв’язуванням рекомбінації перемикання класів (CSR). Рекомбінація перемикання класів — це біологічний механізм, який дозволяє змінювати клас антитіл, що виробляються активованою В-клітиною. Під час CSR частини локусу важкого ланцюга антитіла видаляються з хромосоми, а сегменти гена, що оточують видалену частину, знову з’єднуються, щоб зберегти функціональний ген антитіла, який виробляє антитіло іншого ізотипу. Дволанцюгові розриви ДНК відбуваються на консервативних нуклеотидних мотивах, які називаються перемикаючими (S)-ділянками, які знаходяться вище по ходу від сегментів генів, які кодують константні ділянки важких ланцюгів антитіл. Вони зустрічаються поруч з усіма генами константної ділянки важкого ланцюга, за винятком δ-ланцюга. ДНК розривається на двох обраних S-ділянках через активність низки ферментів, у тому числі цитидин-деамінази (AID), урацил ДНК глікозилази і апіримідин/апуринових (АР)-ендонуклеаз.[5][6] Проміжна ДНК між S-ділянками згодом видаляється з хромосоми, із непотрібними екзонами константної ділянки важкого ланцюга μ або δ, що дозволяє замінити сегмент гена константної області γ, α або ε. Вільні кінці ДНК знову з’єднуються за допомогою процесу, який називається негомологічним з’єднанням кінців (NHEJ), щоб зв’язати екзон варіабельного домену з необхідним нижчим екзоном постійного домену важкого ланцюга антитіла.[7] За відсутності негомологічного з’єднання кінців вільні кінці ДНК можуть бути повторно з’єднані альтернативним шляхом, зміщеним у бік мікрогомологічних з’єднань.[8] За винятком генів μ і δ, В-клітина в будь-який момент часу експресує лише один клас антитіл. Хоча рекомбінація перемикання класів є переважно процесом делеції, перебудовою хромосоми в «цис» позиції, вона також може відбуватися (у 10-20% випадків, залежно від класу Ig) як міжхромосомна транслокація, що змішує гени важкого ланцюга імуноглобуліну з обох алелів.[9][10]
Цитокіни, відповідальні за перемикання класів
Цитокіни Т-клітин модулюють перемикання класів у миші (Таблиця 1) і людини (Таблиця 2).[11][12] Ці цитокіни можуть пригнічувати вироблення IgM.
Регуляторні послідовності генів, відповідальні за перемикання класів
На додаток до надповторюваної структури цільових S-ділянок, процес перемикання класів потребує, щоб S-області спочатку були транскрибовані та сплайсовані з транскриптів важкого ланцюга імуноглобуліну (де вони лежать всередині інтронів). Ремоделювання хроматину, доступність до транскрипції та AID та з'єднання розірваних S-ділянок знаходяться під контролем великого супер-енхансера, розташованого нижче за більш дистальним геном Calpha, 3'-регуляторної області (3'RR).[13] У деяких випадках супер-енхансер 3'RR сам може бути націлений на AID і зазнає розривів ДНК і з’єднання з Sμ, який потім видаляє локус важкого ланцюга Ig і визначає локус суїцидальної рекомбінації (LSR).[14]
| Т-клітини | Цитокіни | Класи імуноглобулінів | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2a | IgG2b | IgG3 | IgG4 | IgE | ||
| Тh2 | ІЛ-4 | ↑ | ↓ | ↓ | ↓ | ↓ | ↑ |
| ІЛ-5 | ↑ | ||||||
| Тh1 | IFNγ | ↓ | ↑ | ↓ | ↑ | ↓ | ↓ |
| Treg | TGFβ | ↑ | ↓ | ↑ | |||
| ІЛ-10 [15] | ↑ | ||||||
| Т-клітини | Цитокіни | Класи імуноглобулінів | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2 | IgG3 | IgG4 | IgA | IgE | ||
| Тh2 | ІЛ-4 | ↑ | ↓ | ↑ | ↑ | ||
| ІЛ-5 | ↑ | ||||||
| Тh1 | IFNγ | ↓ | ↑ | ↓ | |||
| Treg | TGFβ | ↓ | ↑ | ||||
| ІЛ-10 [16][17] | ↑ | ↑ | |||||
Дивіться також
- Антитіла
- Генетична рекомбінація
- Імунний чекпойнт
- Імуногенетика
Посилання
- Bauer G (2020). The variability of the serological response to SARS-corona virus-2: Potential resolution of ambiguity through determination of avidity (functional affinity). Journal of Medical Virology. PMC 7361859. PMID 32633840. doi:10.1002/jmv.26262.
- Eleonora Market, F. Nina Papavasiliou (2003) V(D)J Recombination and the Evolution of the Adaptive Immune System Архівовано 16 лютого 2008 у Wayback Machine. PLoS Biology1(1): e16.
- Evolution of isotype switching. Semin. Immunol. 16 (4): 257–75. 2004. PMID 15522624. doi:10.1016/j.smim.2004.08.005. Проігноровано невідомий параметр
|vauthors=(довідка) - Parham, Peter (2015). The Immune System (вид. 4th). Garland Science.
- Durandy A (2003). Activation-induced cytidine deaminase: a dual role in class-switch recombination and somatic hypermutation. Eur. J. Immunol. 33 (8): 2069–73. PMID 12884279. doi:10.1002/eji.200324133.
- Class switching and Myc translocation: how does DNA break?. Nat. Immunol. 5 (11): 1101–3. 2004. PMC 4625794. PMID 15496946. doi:10.1038/ni1104-1101. Проігноровано невідомий параметр
|vauthors=(довідка) - Roles of nonhomologous DNA end-joining, V(D)J recombination, and class switch recombination in chromosomal translocations. DNA Repair (Amst.) 5 (9–10): 1234–45. 2006. PMID 16793349. doi:10.1016/j.dnarep.2006.05.013. Проігноровано невідомий параметр
|vauthors=(довідка) - IgH class switching and translocations use a robust non-classical end-joining pathway. Nature 449 (7161): 478–82. 2007. PMID 17713479. doi:10.1038/nature06020. Проігноровано невідомий параметр
|vauthors=(довідка) - Interallelic class switch recombination contributes significantly to class switching in mouse B cells. Journal of Immunology 174 (10): 6176–83. May 2005. PMID 15879114. doi:10.4049/jimmunol.174.10.6176. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Immunoglobulin genes undergo legitimate repair in human B cells not only after cis- but also frequent trans-class switch recombination. Genes and Immunity 15 (5): 341–6. 2014. PMID 24848929. doi:10.1038/gene.2014.25. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Janeway CA Jr.; Travers P; Walport M; Shlomchik MJ (2001). Immunobiology. (вид. 5th). Garland Publishing. ISBN 978-0-8153-3642-6. (via NCBI Bookshelf).
- Male D, Brostoff J, Roth DB, Roitt I (2006). Immunology, 7th ed. Philadelphia: Mosby Elsevier, ISBN 978-0-323-03399-2 (pbk.)
- The IgH locus 3' regulatory region: pulling the strings from behind. Advances in Immunology 110. 2011. с. 27–70. ISBN 9780123876638. PMID 21762815. doi:10.1016/B978-0-12-387663-8.00002-8.
- AID-driven deletion causes immunoglobulin heavy chain locus suicide recombination in B cells. Science 336 (6083): 931–4. May 2012. PMID 22539552. doi:10.1126/science.1218692. Проігноровано невідомий параметр
|vauthors=(довідка) - Shparago, N.; Zelazowski, P.; Jin, L.; McIntyre, T. M.; Stuber, E.; Peçanha, L. M.; Kehry, M. R.; Mond, J. J. та ін. (1 травня 1996). IL-10 selectively regulates murine Ig isotype switching. International Immunology 8 (5): 781–790. ISSN 0953-8178. PMID 8671667. doi:10.1093/intimm/8.5.781. Проігноровано невідомий параметр
|doi-access=(довідка); - Human interleukin 10 induces naive surface immunoglobulin D+ (sIgD+) B cells to secrete IgG1 and IgG3. The Journal of Experimental Medicine 179 (2): 757–62. February 1994. PMC 2191366. PMID 8294883. doi:10.1084/jem.179.2.757. Проігноровано невідомий параметр
|vauthors=(довідка) - Interleukin-10 induces immunoglobulin G isotype switch recombination in human CD40-activated naive B lymphocytes. The Journal of Experimental Medicine 183 (3): 937–47. March 1996. PMC 2192363. PMID 8642297. doi:10.1084/jem.183.3.937. Проігноровано невідомий параметр
|vauthors=(довідка)