B-клітинний рецептор
В-клітинний рецептор (BCR) — це трансмембранний білок на поверхні В-клітини. BCR складається з молекули імуноглобуліну, зв’язаної з мембраною, і фрагмента, що передає сигнал. Перший утворює трансмембранний рецепторний білок 1 типу і зазвичай розташований на зовнішній поверхні цих лімфоцитів.[1] За допомогою біохімічної передачі сигналів і фізичного контакту з антигеном (імунний синапс) BCR контролює активацію В-клітини.[2] В-клітини здатні збирати і захоплювати антигени, залучаючи біохімічні модулі для кластеризації рецепторів, розмножуватись, створювати сили тяжіння і транспортувати рецептори, що в кінцевому підсумку завершується ендоцитозом і презентацією антигена.[1] Механічна активність В-клітин дотримується схеми негативних і позитивних зворотних зв'язків, які регулюють кількість видаленого антигена шляхом безпосереднього маніпулювання динамікою зв'язку BCR-антиген.[3] Зокрема, групування та розповсюдження антигена посилює зв’язок антиген-BCR, тим самим посилюючи його чутливість та ампліфікацію.[4] З іншого боку, сили тяжіння відокремлюють антиген від BCR, таким чином перевіряючи силу зв'язку.

Зв’язувальна частина BCR складається з мембранозв’язаного антитіла, яке, як і всі антитіла, має два ідентичних паратопи, які є унікальними та визначаються випадковим чином. BCR до антигена є важливим датчиком, який необхідний для активації, виживання та розвитку В-клітин. В-клітина активується під час першої зустрічі з антигеном (його «спорідненим антигеном»), який зв’язується з його рецептором, що призводить до проліферації та диференціювання клітин для створення популяції В-клітин плазми, які секретують антитіла, і В-клітин пам’яті.[1][4] BCR виконує дві важливі функції при взаємодії з антигеном. Однією з функцій є передача сигналу, що включає зміни в олігомеризації рецепторів.[1] Друга функція полягає в опосередковуванні інтерналізації для подальшої обробки антигену та презентації пептидів Т-хелперам.
Розвиток і структура В-клітинного рецептора
Першою контрольною точкою у розвитку В-клітини є вироблення функціонального пре-BCR, який складається з двох сурогатних легких ланцюгів і двох важких ланцюгів імуноглобуліну, які зазвичай пов’язані з сигнальними молекулами Ig-α (або CD79A) і Ig-β (або CD79B).[1][5] Кожна В-клітина, що виробляється в кістковому мозку, є високоспецифічною до конкретного антигена.[1][3] BCR можна знайти в ряді ідентичних копій мембранних білків, які експонуються на поверхні клітини.[1][3][6]
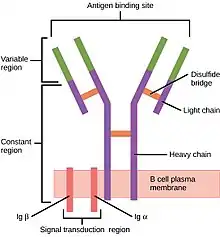
В-клітинний рецептор складається з двох частин:
- Зв’язана з мембраною молекула імуноглобуліну одного ізотипу (IgD, IgM, IgA, IgG або IgE). За винятком наявності цілісного мембранного домену, вони ідентичні мономерній версії своїх секретованих форм.
- Ділянка передачі сигналу: гетеродимер під назвою Ig-α/Ig-β (CD79), зв’язаний між собою дисульфідними містками. Кожен член димеру вставлений у плазматичну мембрану і має цитоплазматичний хвіст, що несе мотив активації імунорецепторів на основі тирозину (ITAM).[6][7]
Більш аналітично, комплекс BCR складається з антигензв’язувальної субодиниці, відомої як мембранний імуноглобулін (mIg), яка складається з двох легких ланцюгів імуноглобуліну (IgL) і двох важких ланцюгів імуноглобуліну (IgHs), а також двох гетеродимерних субодиниць Ig- α і Ig-β. Для того щоб молекули mIgM могли транспортуватися на поверхню клітини, має бути поєднання Ig-α та Ig-β з молекулами mIgM. Пре-В-клітини, які не генерують жодної молекули Ig, зазвичай на своїй поверхні несуть як Ig-α, так і Ig-β.[1][7]
Гетеродимери можуть існувати в В-клітинах у вигляді асоціації або комбінації з іншими специфічними для В-клітин білками або окремо, замінюючи таким чином молекулу mIgM. У BCR частина, яка розпізнає антигени, складається з трьох різних ділянок, які називаються V, D і J. [1][4][8] Існує ряд генів, які кодують кожну з цих ділянок в геномі, і які можуть бути рекомбіновані та об’єднані різними способами для створення широкого спектру рецепторних молекул.[1][4][7][8] Виробництво цього спектру має вирішальне значення, оскільки організм може зустріти набагато більше антигенів, ніж має генів. За допомогою цього процесу організм має спосіб утворення кількох різних комбінацій рецепторних молекул, що розпізнають антиген. Перебудова важкого ланцюга BCR супроводжує початкові кроки в розвитку В-клітини. Короткі області JH (joining - з’єднання) і DH (diversity - різноманіття) спочатку рекомбінуються в ранніх про-В-клітинах у процесі, який залежить від ферментів RAG2 і RAG1.[8][9] Після рекомбінації D і J-областей, клітина тепер називається «пізньою про-В» клітиною, а коротка ділянка DJ тепер може бути рекомбінована з довшим сегментом гена VH.[7][8]
BCR мають певні сайти зв’язування, які забезпечвють комплементарність поверхні епітопу та поверхні рецептора, часто за рахунок нековалентних сил. За відсутності специфічного антигена, зрілі В-клітини можуть виживати в периферійному кровообігу лише обмежений час. Це тому, що, коли клітини не зустрічають жодного антигена протягом цього часу, в них запуститься апоптоз.[6] Примітно, що в периферичній циркуляції апоптоз важливий для підтримки оптимального обігу В-лімфоцитів.[8][9] За структурою BCR до антигенів майже ідентичні вільним антитілам.[1][5] Однак у С-кінцевій ділянці важких ланцюгів є характерна структурна відмінність, яка складається з короткого гідрофобного відрізка, який перетинає ліпідний бішар мембрани.
Сигнальні шляхи В-клітинного рецептора
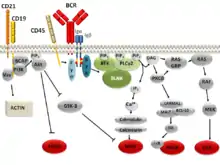
Існує кілька сигнальних шляхів BCR, які ініціюються, коли субодиниці mIg BCR зв’язують специфічний антиген. Початкова активація BCR подібна для всіх рецепторів сімейства некаталітичних тирозинофосфорильованих рецепторів.[11] Зв'язування дозволяє фосфорилювати мотиви активації імунорецепторів на основі тирозину (ITAM) в асоційованих субодиницях гетеродимеру Igα/Igβ тирозинкіназами сімейства Src, включаючи Blk, Lyn і Fyn. Було запропоновано кілька моделей того, як зв’язування BCR-антиген індукує фосфорилювання, включаючи конформаційну зміну рецептора та агрегацію кількох рецепторів при зв’язуванні антигена.[12] Тирозинкіназа Syk зв’язується і активується фосфорильованими ITAM і, у свою чергу, фосфорилює каркасний білок BLNK на кількох сайтах. Після фосфорилювання низхідні сигнальні молекули залучаються до BLNK, що призводить до їх активації та передачі сигналу всередину.[13]
- Шлях фактора транскрипції IKK/NF-κB: CD79 та інші білки, мікросигналосоми, активують PLC-γ після розпізнавання антигену BCR і до того, як він почне зв'язуватися з c-SMAC. Потім він розщеплює PIP2 на IP3 і DAG (діацилгліцерин). IP3 діє як вторинний месенджер різкого збільшення іонного кальцію всередині цитозолю (через вивільнення з ендоплазматичного ретикулума або надходження із позаклітинного середовища через іонні канали). Це призводить до кінцевої активації PKCβ з кальцію та DAG. PKCβ фосфорилює (прямо або опосередковано) білок сигнального комплексу NF-κB CARMA1 (сам комплекс, що містить CARMA1, BCL10 і MALT1). Це призводить до залучення та виклику IKK (кінази IkB), TAK1, кількома ферментами убіквітилювання, також пов’язаними з комплексом CARMA1/BCL10/MALT1. Сам MALT1 є каспазоподібним білком, який розщеплює А20, інгібує білок сигналізації NF-kB (який діє яе деубіквітатор убіквітованих субстратів NF-кВ, маючи інгібуючий ефект). TAK1 фосфорилює тример IKK після того, як він також був залучений до сигнального комплексу за допомогою пов’язаних з ним ферментів убіквітилювання. Потім IKK фосфорилює IkB (інгібітор NF-κB і зв’язаний з ним), що викликає його руйнування, позначаючи його для протеолітичної деградації, звільняючи цитозольний NF-κB. Потім NF-κB мігрує в ядро, щоб зв’язатися з ДНК на специфічних елементах відповіді, викликаючи рекрутування транскрипційних молекул і розпочинаючи процес транскрипції.
- Зв’язування ліганда з BCR також призводить до фосфорилювання білка BCAP. Це призводить до зв’язування та активації кількох білків з доменами SH2, що зв’язують фосфотирозин. Одним з таких білків є PI3K. Активація PI3K призводить до фосфорилювання PIP2, утворюючи PIP3. Білки з доменами PH (гомологія плекстрину) можуть зв’язуватися з щойно створеним PIP3 і активуватися. До них відносяться білки сімейства FoxO, які стимулюють прогресування клітинного циклу, і протеїнкіназа D, яка посилює метаболізм глюкози. Іншим важливим білком з доменом PH є Bam32. Це залучає та активує невеликі GTPaze, такі як Rac1 і Cdc42. Вони, у свою чергу, відповідають за зміни цитоскелета, пов’язані з активацією BCR шляхом модифікації полімеризації актину.
В-клітинний рецептор при злоякісних пухлинах
Було показано, що В-клітинний рецептор бере участь у патогенезі різних В-клітинних лімфоїдних новоутворень. Хоча можливо, що стимуляція зв’язуванням антигена сприяє проліферації злоякісних В-клітин[14], все більше доказів свідчить про те, що антигеннезалежна самоактивація BCR є ключовою ознакою зростаючої кількості В-клітинних неоплазій [15][16][17][18] В даний час передача сигналів рецептором В-клітин є терапевтичною мішенню при різних лімфоїдних новоутвореннях.[19]
Дивіться також
Посилання
- Owen, J.; Punt, J.; Stranford, S; Jones, P.; Kuby, J. (2013). Kuby Immunology (Seventh ed.). New York: W.H. Freeman and Company. с. 102–104. ISBN 978-1429219198.
- Saito, Batista; Saito, Takashi; Facundo, D. (2010). Immunological Synapse (Current Topics in Microbiology and Immunology, 340). Berlin: Heidelberg: Springer Berlin Heidelberg. ISBN 978-3642038570.
- Merlo, Lauren M. F.; Mandik-Nayak, Laura (2013). Cancer Immunotherapy: Chapter 3-Adaptive Immunity: B Cells and Antibodies.. London: Academic Press; 2 edition. с. 25–40. ISBN 978-0-12-394296-8.
- Dal Porto, JM; Gauld, SB (2014). B cell antigen receptor signaling 101. Merrell KT, Mills D, Pugh-Bernard AE. Mol Immunol 41: 599–613. PMID 15219998. doi:10.1016/j.molimm.2004.04.008.
- Brenzski, Randall J.; Monroe, John G. (2010). Chapter 2: B cell Receptor. У Sigalov, Alexander B. Multichain Immune Recognition Receptor Signaling: From Spatiotemporal Organization to Human Disease (Advances in Experimental Medicine and Biology). Springer; Softcover reprint of hardcover 1st ed. 2008 edition (November 23, 2010). с. 12–21. ISBN 978-1441918871.
- Janeway, CA Jr; Travers, P.; Walport, M. (2015). Immunobiology: The Immune System in Health and Disease. New York: Garland Science (5th edition). ISBN 978-0815341017.
- Pier, Gerland B.; Lyczak, Jeffrey B.; Wetzler, Lee M. (2005). Immunology, Infection, and Immunity. Washington D.C.: ASM Press. с. 234–247. ISBN 978-1555812461.
- Hoehn, Kenneth B.; Fowler, Anna; Lunter, Gerton; Pybus, Oliver G. (2016). The Diversity and Molecular Evolution of B Cell Receptors during Infection. Mol Biol Evol 33: 1147–57. PMC 4839220. PMID 26802217. doi:10.1093/molbev/msw015.
- Alberts, Bruce (2014). Molecular Biology of the Cell. Garland Science; 6th edition. ISBN 978-0815344322.
- Wan, Leo D.; Clark, Marcus R. (2003). B cell antigen-receptor signaling in lymphocyte development. Immunology 110: 411–20. PMC 1783068. PMID 14632637. doi:10.1111/j.1365-2567.2003.01756.x.
- Non-catalytic tyrosine- phosphorylated receptors. Immunological Reviews 250 (1): 258–276. November 2012. PMID 23046135. doi:10.1111/imr.12008. Проігноровано невідомий параметр
|vauthors=(довідка) - B cell receptor: from resting state to activated. Immunology 136 (1): 21–27. 2012. PMC 3372753. PMID 22269039. doi:10.1111/j.1365-2567.2012.03564.x. Проігноровано невідомий параметр
|vauthors=(довідка) - Janeway's immunobiology (вид. 8th). Garland Science. 2011. с. 258 – 260. ISBN 0815342438.
- Leukemia and auto-immunization- some possible relationships. Blood 14: 1151–8. 1959. PMID 13813891. Проігноровано невідомий параметр
|vauthors=(довідка) - Oncogenic potential of the B cell antigen receptor and its relevance to heavy chain diseases and other B-cell neoplasias: a new model. Research in Immunology 141 (6): 543–53. 1990. PMID 2284498. doi:10.1016/0923-2494(90)90022-Q. Проігноровано невідомий параметр
|vauthors=(довідка) - Pre-B-cell development in the absence of lambda 5 in transgenic mice expressing a heavy-chain disease protein. Current Biology 5 (10): 1140–8. October 1995. PMID 8548286. doi:10.1016/S0960-9822(95)00230-2. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Chronic active B-cell-receptor signalling in diffuse large B-cell lymphoma. Nature 463 (7277): 88–92. January 2010. Bibcode:2010Natur.463...88D. PMC 2845535. PMID 20054396. doi:10.1038/nature08638. Проігноровано невідомий параметр
|vauthors=(довідка) - Chronic lymphocytic leukaemia is driven by antigen-independent cell-autonomous signalling. Nature 489 (7415): 309–12. September 2012. Bibcode:2012Natur.489..309M. PMID 22885698. doi:10.1038/nature11309. Проігноровано невідомий параметр
|vauthors=(довідка) - The B-cell receptor signaling pathway as a therapeutic target in CLL. Blood 120 (6): 1175–84. August 2012. PMC 3418714. PMID 22715122. doi:10.1182/blood-2012-02-362624. Проігноровано невідомий параметр
|vauthors=(довідка)
