Імуноглобуліни класу A
Імуноглобуліни класу A (IgA, також sIgA в секреторній формі) — антитіло, яке відіграє вирішальну роль в імунній функції слизових оболонок. Кількість IgA, що виробляється на мембранах слизових оболонок, перевищує всі інші типи антитіл у поєднанні.[3] В абсолютному вираженні, від трьох до п'яти грамів IgA секретируются в просвіт кишечника кожен день.[4] Це становить до 15 % загальної кількості імуноглобулінів, що виробляються в організмі.[5]
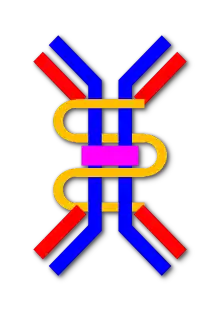
IgA має два підкласи (IgA1 та IgA2) і може вироблятися як у мономерній, так і в димерній формі. Димерна форма IgA є найрозповсюдженішою, її також називають секреторною IgA (sIgA). sIgA — основний імуноглобулін, що міститься у слизових секретах, включаючи сльози, слину, піт, молозиво та секрети сечостатевих шляхів, шлунково-кишкового тракту, простати та епітелію дихальних шляхів. Він також міститься в невеликій кількості в крові. Секреторна складова sIgA захищає імуноглобулін від руйнування протеолітичними ферментами; таким чином, sIgA може вижити в суворих умовах шлунково-кишкового тракту і забезпечити захист від мікробів, які розмножуються в секретах організму.[6] sIgA також може пригнічувати запальний ефект інших імуноглобулінів.[7] IgA є поганим активатором системи комплементу, і опсонізується лише слабко.
Форми
IgA1 та IgA2
IgA існує у двох ізотипах — IgA1 та IgA2. Вони обидва сильно глікозильовані білки.[8] Хоча IgA1 переважає у сироватці крові (~80 %), відсоток IgA2 у секретах вищий, ніж у сироватці крові (~35 % у секретах);[9] співвідношення клітин, що секретують IgA1 та IgA2, різниться в різних лімфоїдних тканинах людського тіла:[10]
- IgA1 — це переважний підклас IgA, який міститься в сироватці крові. У більшості лімфоїдних тканин переважання клітини, що виробляють IgA1.[11]
- У IgA2 важкі та легкі ланцюги пов'язані не дисульфідом, а нековалентними зв'язками. У секреторних лімфоїдних тканинах (наприклад, лімфоїдна тканина, асоційована з кишківником або GALT) частка вироблення IgA2 більша, ніж у несекреторних лімфоїдних органах (наприклад, селезінці, периферичних лімфатичних вузлах).
Як IgA1, так і IgA2 виявлені у зовнішніх секретах, таких як молозиво, материнське молоко, сльози та слина, де IgA2 є більш помітним, ніж у крові.[9] Полісахаридні антигени, як правило, індукують більше IgA2, ніж білкові антигени.[10]
І IgA1, і IgA2 можуть бути у мембранно-зв'язаній формі.[12] (див. B-клітинний рецептор)
Сироватковий та секреторний IgA
Також можна розрізнити форми IgA за їх місцем розташування — сироватковий IgA nf секреторнbq IgA.
У секреторному IgA, формі, що міститься в секретах, полімери 2–4 мономерів IgA пов'язані двома додатковими ланцюгами; як така, молекулярна маса slgA становить 385 000 а.о.м. Одним з них є J-ланцюг (приєднувальний ланцюг), який є поліпептидом молекулярної маси 15 000 а.о.м., багатим цистеїном і структурно повністю відмінним від інших ланцюгів імуноглобулінів. Цей ланцюг утворюється в клітинах, що секретують IgA.
Олігомерні форми IgA у зовнішніх (слизових) секретах також містять поліпептид набагато більшої молекулярної маси (70 000 а.о.м.), який називається секреторною складовою, що виробляється епітеліальними клітинами. Ця молекула походить від рецептора полі-Ig (130 000 а.о.м.), який відповідає за поглинання та трансцелюлярне транспортування олігомерного (але не мономерного) IgA через епітеліальні клітини та у виділення, такі як сльози, слина, піт та кишкова рідина.
Фізіологія
Сироватковий IgA
У крові IgA взаємодіє з Fc-рецептором, що називається FcαRI (або CD89), який експресується на імунних ефекторних клітинах, ініціюючи запальні реакції.[13] Лігування FcαRI IgA, що містить імунні комплекси, спричиняє антитілозалежну клітинно-опосередковану цитотоксичність (АЗКЦ), дегрануляцію еозинофілів та базофілів, фагоцитоз моноцитами, макрофагами та нейтрофілами та активацію респіраторної активності поліморфно-ядерних лейкоцитів.[13]
Секреторний IgA
Висока концентрація IgA на слизових ділянках є результатом взаємодії між плазматичними клітинами, що виробляють полімерний IgA (pIgA), і клітинами епітелію слизової, які експресують рецептор імуноглобуліну, який називається полімерним рецептором імуноглобулінку (pIgR).[13] Полімерний IgA (головним чином секреторний димер) виробляється плазматичними клітинами у власній пластинці, прилеглої до поверхонь слизової. Він зв'язується з полімерним рецептором імуноглобуліну на базолатеральній поверхні епітеліальних клітин і потрапляє в клітину шляхом ендоцитозу. Комплекс рецептор-IgA проходить через клітинні відділи, перш ніж виділятися на просвітню поверхню епітеліальних клітин, все ще приєднану до рецептора. Відбувається протеоліз рецептора, і димерна молекула IgA, разом з частиною рецептора, секреторною складовою, відомою як sIgA, можуть вільно дифундувати по всьому просвіту.[14] У кишківнику IgA може зв'язуватися із шаром слизу, що покриває епітеліальні клітини. Таким чином формується бар'єр, здатний нейтралізувати загрози до того, як вони досягнуть епітеліальних клітин.
Утворення sIgA проти специфічних антигенів залежить від відбору зразків М-клітин і основних дендритних клітин, активації Т-клітин та переходу класу В-клітин у GALT, брижових лімфатичних вузлів та ізольованих лімфоїдних фолікул у тонкому кишечнику.[15]
sIgA в основному діє шляхом блокування епітеліальних рецепторів (наприклад, шляхом зв'язування їх лігандів із патогенними мікроорганізмами), стерично перешкоджаючи прикріпленню до клітин епітелію та імунної ексклюзії.[15] Імунна ексклюзія — це процес аглютинації полівалентних антигенів або патогенів, зшиваючи їх з антитілами, захоплюючи в шар слизу та/або очищаючи перистальтично. Олігосахаридні ланцюги компонента IgA можуть поєднуватися зі слизовим шаром, який розміщений над епітеліальними клітинами.[15] Оскільки sIgA є поганим опсоніном і активатором комплементу, простого зв'язування патогена не обов'язково достатньо для його утримання — певні епітопи, можливо, повинні бути пов'язані, щоб стерично перешкоджати доступу до епітелію.[15]
Кліренс IgA принаймні частково опосередковується азіалоглікопротеїновими рецепторами, які розпізнають N-глікани IgA, що припиняються галактозою.[8]
Патологія
Генетична
Зниження або відсутність IgA через спадкову нездатність виробляти IgA називається селективним дефіцитом імуноглобулінів класу А і може спричинити клінічно значущий імунодефіцит.[16]
Антитіла до IgA, іноді присутні у осіб з низьким рівнем або відсутністю IgA, можуть спричинити серйозні анафілактичні реакції при переливанні продуктів крові, які випадково містять IgA. Однак більшість людей із підозрою на анафілактичні реакції до IgA мали гострі генералізовані реакції, спричинені іншими чинниками, крім переливання продуктів крові із IgA.[17]
Мікробна
Види Neisseria, включаючи Neisseria gonorrhoeae (що викликає гонорею),[18] Streptococcus pneumoniae[19] та Haemophilus influenzae типу B,[20] всі виділяють протеазу, яка руйнує IgA. Крім того, було доведено, що види Blastocystis мають кілька підтипів, що утворюють ферменти цистеїнової та аспарагінової протеази, які розкладають (деградуються) IgA людини.[21]
Аутоімунні та імунно-опосередковані
IgA-нефропатія спричиняється відкладеннями IgA в нирках. Поки невідомо, чому відкладення IgA виникають при цьому хронічному захворюванні. Деякі теорії припускають, що аномалії імунної системи призводять до цих відкладень.[22]
Целіакія включає патологію IgA через наявність антиендомизиальних антитіл IgA.[23][24]
Геморагічний васкуліт — це системний васкуліт, спричинений відкладеннями IgA та компонента 3 системи комплементу (С3) у дрібних судинах. Геморагічний васкуліт зазвичай виникає у маленьких дітей і вражає шкіру та сполучні тканини, мошонку, суглоби, шлунково-кишковий тракт та нирки. Зазвичай це стає ускладенняи після інфекцій верхніх дихальних шляхів і розсмоктується протягом декількох тижнів, коли печінка очищає сукупності IgA.[25]
Лінійний бульозний дерматоз IgA та пемфігус IgA — два приклади імунобульозних захворювань, опосередкованих IgA. Імунобульозні захворювання, опосередковані IgA, часто важко піддаються лікуванню навіть за допомогою зазвичай ефективних препаратів, таких як ритуксимаб.[26]
Індукована ліками
Ванкоміцин може індукувати лінійний бульозний дерматоз IgA у деяких пацієнтів.[27]
Примітки
- А. Боннер, А. Альмогрен, П. Б. Фуртадо, М. А. Керр, С. Дж. Перкінс (Січень 2009). Location of secretory component on the Fc edge of dimeric IgA1 reveals insight into the role of secretory IgA1 in mucosal immunity. Mucosal Immunology 2 (1): 74–84. PMID 19079336. doi:10.1038/mi.2008.68.
- А. Боннер, А. Альмогрен, П. Б. Фуртадо, М. А. Керр, С. Дж. Перкінс (Лютий 2009). The nonplanar secretory IgA2 and near planar secretory IgA1 solution structures rationalize their different mucosal immune responses. The Journal of Biological Chemistry 284 (8): 5077–87. PMC 2643523. PMID 19109255. doi:10.1074/jbc.M807529200.
- С. Фагарасан, Т. Гонджо (Січень 2003). Intestinal IgA synthesis: regulation of front-line body defences. Nature Reviews. Immunology 3 (1): 63–72. PMID 12511876. doi:10.1038/nri982.
- П. Брандцаг, Р. Пабст (Листопад 2004). Let's go mucosal: communication on slippery ground. Trends in Immunology 25 (11): 570–7. PMID 15489184. doi:10.1016/j.it.2004.09.005.
- А. Дж. Макферсон, Е. Слак (Листопад 2007). The functional interactions of commensal bacteria with intestinal secretory IgA. Current Opinion in Gastroenterology 23 (6): 673–8. PMID 17906446. doi:10.1097/MOG.0b013e3282f0d012.
- Луїс К. Жункейра; Хосе Карнейро (2003). Basic Histology. McGraw-Hill. ISBN 978-0-8385-0590-8.
- Дж. Голмгрен, Ч. Черкінскі (Квітень 2005). Mucosal immunity and vaccines. Nature Medicine 11 (4 Suppl): S45–53. PMID 15812489. doi:10.1038/nm1213.
- Е. Маверакіс, К. Кім, М. Шимода, М. Є. Гершвін, Ф. Патель, Р. Вілкен, С. Райчаудхурі, Л. Р. Рухаак, К. Б. Лебрілла (Лютий 2015). Glycans in the immune system and The Altered Glycan Theory of Autoimmunity: a critical review. Journal of Autoimmunity 57: 1–13. PMC 4340844. PMID 25578468. doi:10.1016/j.jaut.2014.12.002.
- Д. Л. Делакруа, К. Дайв, Дж. К. Рамбо, Дж. П. Ваерман (Жовтень 1982). IgA subclasses in various secretions and in serum. Immunology 47 (2): 383–5. PMC 1555453. PMID 7118169.
- Б. Сімелл, Т. Кілпі, Г. Кайгти (Березень 2006). Subclass distribution of natural salivary IgA antibodies against pneumococcal capsular polysaccharide of type 14 and pneumococcal surface adhesin A (PsaA) in children. Clinical and Experimental Immunology 143 (3): 543–9. PMC 1809616. PMID 16487254. doi:10.1111/j.1365-2249.2006.03009.x.
- А. Дж. Макферсон, К. Д. Маккой, Ф. Е. Йохансен, П. Брандцаг (Січень 2008). The immune geography of IgA induction and function. Mucosal Immunology 1 (1): 11–22. PMID 19079156. doi:10.1038/mi.2007.6.
- А. Ф. Хунг, Дж. Б. Чень, Т. В. Чанг (Серпень 2008). Alleles and isoforms of human membrane-bound IgA1. Molecular Immunology 45 (13): 3624–30. PMID 18538846. doi:10.1016/j.molimm.2008.04.023.
- В. Сноек, І. Р. Пітерс, Е. Кокс (2006). The IgA system: a comparison of structure and function in different species. Veterinary Research 37 (3): 455–67. PMID 16611558. doi:10.1051/vetres:2006010.
- К. С. Кецель, Дж. К. Робінсон, К. Р. Чінталачаруву, Дж. П. Ваерман, М. Е. Ламм (Жовтень 1991). The polymeric immunoglobulin receptor (secretory component) mediates transport of immune complexes across epithelial cells: a local defense function for IgA. Proceedings of the National Academy of Sciences of the United States of America 88 (19): 8796–800. Bibcode:1991PNAS...88.8796K. PMC 52597. PMID 1924341. doi:10.1073/pnas.88.19.8796.
- Н. Дж. Мантіс, Н. Рол, Б. Кортезі (Листопад 2011). Secretory IgA's complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunology 4 (6): 603–11. PMC 3774538. PMID 21975936. doi:10.1038/mi.2011.41.
- Л. Єль (Січень 2010). Selective IgA deficiency. Journal of Clinical Immunology 30 (1): 10–6. PMC 2821513. PMID 20101521. doi:10.1007/s10875-009-9357-x.
- С. Г. Сендлер, Д. Меллорі, Д. Маламут, Р. Екріч (Січень 1995). IgA anaphylactic transfusion reactions. Transfusion Medicine Reviews 9 (1): 1–8. PMID 7719037. doi:10.1016/S0887-7963(05)80026-4.
- Р. Галтер, Дж. Полнер, Т. Ф. Мейєр (Липень 1984). IgA protease of Neisseria gonorrhoeae: isolation and characterization of the gene and its extracellular product. The EMBO Journal 3 (7): 1595–601. PMC 557564. PMID 6430698. doi:10.1002/j.1460-2075.1984.tb02016.x.
- М. Проктор, П. Дж. Меннінг (Вересень 1990). Production of immunoglobulin A protease by Streptococcus pneumoniae from animals. Infection and Immunity 58 (9): 2733–7. PMC 313560. PMID 2117567. doi:10.1128/IAI.58.9.2733-2737.1990.
- Дж. В. Ст. Джейм, М. Л. де ла Морена, С. Фалков (Жовтень 1994). A Haemophilus influenzae IgA protease-like protein promotes intimate interaction with human epithelial cells. Molecular Microbiology 14 (2): 217–33. PMID 7830568. doi:10.1111/j.1365-2958.1994.tb01283.x.
- Т. Робертс, Д. Старк, Дж. Гаркнесс, Дж. Елліс (2014). Update on the pathogenic potential and treatment options for Blastocystis sp. Gut Pathogens 6: 17. PMC 4039988. PMID 24883113. doi:10.1186/1757-4749-6-17.
- article/239927 on eMedicine
- Г. Е. Принс, Г. Л. Норман, В. Л. Біндер (Березень 2000). Immunoglobulin A (IgA) deficiency and alternative celiac disease-associated antibodies in sera submitted to a reference laboratory for endomysial IgA testing. Clinical and Diagnostic Laboratory Immunology 7 (2): 192–6. PMC 95847. PMID 10702491. doi:10.1128/cdli.7.2.192-196.2000.
- Ш. Каннінгем-Рундлз (Вересень 2001). Physiology of IgA and IgA deficiency. Journal of Clinical Immunology 21 (5): 303–9. PMID 11720003. doi:10.1023/A:1012241117984.
- А. Рай, К. Наст, С. Адлер (December 1999). Henoch-Schönlein purpura nephritis. Journal of the American Society of Nephrology 10 (12): 2637–44. PMID 10589705.
- Й. Гі, М. Шимода, Ю. Оно, І. Б. Віллалобос, А. Мітра, Т. Конія, С. А. Грандо, Дж. Дж. Зона, Е. Маверакіс (June 2015). Persistence of Autoreactive IgA-Secreting B Cells Despite Multiple Immunosuppressive Medications Including Rituximab. JAMA Dermatology 151 (6): 646–50. PMID 25901938. doi:10.1001/jamadermatol.2015.59.
- Дж. Реймонд Го; Омар М. Абу Салех (15 жовтня 2020). Vancomycin-Induced Linear IgA Bullous Dermatosis. New England Journal of Medicine (англ.) 383 (16): 1577–1577. ISSN 0028-4793. doi:10.1056/NEJMicm2003334.