Селеноцистеїн
Селе́ноцистеї́н (скороч. Sec або U, в ранніх публікаціях зустрічається скорочення Se-Cys)[1] — органічна сполука природного походження, 21-ша протеїногенна амінокислота.
| Селеноцистеїн | |
|---|---|
 | |
| Ідентифікатори | |
| Абревіатури | Sec |
| Номер CAS | 10236-58-5 |
| Номер EINECS | 808-428-7 |
| DrugBank | 02345 |
| KEGG | C05688 |
| ChEBI | 16633 |
| SMILES |
O=C(O)[C@@H](N)C[SeH] |
| InChI |
InChI=1S/C3H7NO2Se/c4-2(1-7)3(5)6/h2,7H,1,4H2,(H,5,6)/t2-/m0/s1 |
| Номер Бельштейна | 2498378 |
| Властивості | |
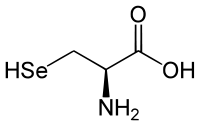
| Молекулярна формула | C3H7NO2Se |
| Молярна маса | 168,05 г/моль |
| Якщо не зазначено інше, дані наведено для речовин у стандартному стані (за 25 °C, 100 кПа) | |
| Інструкція з використання шаблону | |
| Примітки картки | |
Селеноцистеїн та селеноцистеїнові білки зустрічаються в трьох доменах живого.[2] Селеноцистеїн є аналогом цистеїну із селенвмісною хімічною групою –SeH замість сірковмісної тіольної групи –SH.
Селеноцистеїн входить до складу деяких ензимів (в тому числі до складу глутатіонпероксидази, тіоредоксинредуктази, форматдегідрогенази, гліцинредуктази, метіонін-R-сульфоксидредуктази B1 (SEPX1), деяких гідрогеназ тощо).
Селеноцистеїн був відкритий в 1974 році.[3]
Структура
Хімічна будова селеноцистеїну подібна до цистеїну. Єдиною відмінністю між ними є наявність атому селену замість атому сірки. Селенольна група селеноцистеїну є депротонованою за фізіологічних значень pH. Протеїни, що містять один або декілька залишків селеноцистеїну, мають назву селенопротеїни. Більшість селенопротеїнів має лише один залишок селеноцистеїну. Селенопротеїни, що виконують каталітичні функції і чиї каталітичні функції критично залежать від наявності сленоцистеїнового залишку в активному сайті, мають назву селеноензимів.[4]
Біологія
Селеноцистеїн одночасно має нижчий показник pKa (5,47) та нижчий окисно-відновний потенціал порівняно з цистеїном. Ця комбінація властивостей роблять його дуже вдалою заміною цистеїну для білків, що виконують антиоксидантні функції.[5]
Незважаючи на те, що ця амінокислота була ідентифікована в усіх трьох доменах живого, вона не є універсальною для всіх організмів.[6] На відміну від двадцяти інших канонічних протеїногенних амінокислот, селеноцистеїн не кодується напряму (тобто йому однозначно не відповідає жоден з кодонів) в генетичному коді.[7] Селеноцистеїн кодується кодоном UGA в мРНК, тобто кодоном, що зазвичай має значення стоп-кодона і означає кінець трансляції. Цей спосіб розширення канонічного генетичного коду отримав назву «трансляційне рекодування» (англ. translational recoding)[8] Ефективність трансляційного рекодування залежить від конкретного селенопротеїну та від наявності факторів ініціації.[9] Якщо клітини знаходяться в селенодифіцитних умовах, трансляція селенопротеїнів обривається на UGA-кодоні, що дає вкорочений нефункціональний білок. Для встроювання селеноцистеїну у відповідь на кодон UGA необхідна наявність спеціального цис-регуляторного елементу в мРНК селенопротеїну, так званого SECIS-елементу. SECIS-елемент має характерну послідовність нуклеотидів і специфічну шпилькоподібну вторинну структуру. У бактерій SECIS-елемент зазвичай розташований одразу після UGA-кодону в межах відкритої рамки зчитування селенопротеїну.[10] У архей та еукаріот SECIS-елементи зазвичай розташовані в 3'-нетрансльованому регіоні (3' UTR) мРНК, та можуть трансляційно рекодувати одразу декілька UGA-кодонів.[11]
На відміну від канонічних амінокислот, клітини майже не містять селеноцистеїн у вільному стані. Вони зберігають селен у вигляді селеноводню (H2Se). Синтез селеноцистеїну відбувається на спеціальних тРНК, які потім беруть участь у біосинтезі селенопротеїнів.
Первинна та вторинна структура селеноцистеїнової тРНК, тРНКSec, відрізняється від звичайних тРНК. Найбільшими відмінністю є акцепторна ділянка 8-bp (бактерії) або 10-bp (еукаріоти), довга варіабельна ділянка, а також нуклеотидні заміни в деяких консервативних позиціях. Селеноцистеїнова тРНК спочатку ацилюється серином за допомогою серил-тРНК-лігази, але Ser-тРНКSec, яка утворюється, не бере участі в трансляції, тому що вона не розпізнається факторами елонгації трансляції (EF-Tu у бактерій та eEF1A у еукаріот).
Залишок серину, зв'язаний із селеноцистеїновою тРНК, далі перетворюється на селеноцистеїн під дією пірідоксальфосфатвмісного ензиму селеноцистеїнсинтетази. У еукаріот та архей для успішного завершення цього процесу необхідні два ензими: PSTK [O-фосфосерил-тРНК[Ser]Sec-кіназа] та селеноцистеїнсинтетаза.[12][13] Утворена Sec-тРНКSec селективно приєднується до альтернативних факторів елонгації транскрипції (SelB або mSelB (або eEFSec)), які доставляють її на рибосому, на якій відбувається трансляція мРНК селенопротеїну. Селективність механізму доставки забезпечується наявністю додаткового білкового домену (в бактеріях — SelB) або додаткової білкової субодиниці (SBP2 для еукаріотичних mSelB/eEFSec), які споріднені до SECIS-елементів селенопротеїнових мРНК.
Станом на 2003 рік було відомо 25 людських білків, що містять селеноцистеїн.[14]
Селеноцистеїнові поіхідні, γ-глутаміл-Se-метлиселеноцистеїн та Se-метилселеноцистеїн, знайдені в рослинах родів Allium та Brassica.[15]
Застосування
Біотехнологічні застосування селеноцистеїну включають використання 73Se-міченого Sec (час напіврозпаду 73Se складає 7,2 годин) в позитрон-емісійній томографії (PET) та 75Se-міченого Sec (час напіврозпаду 75Se — 118,5 днів) в специфічному радіоміченні, рентгеноструктурному аналізі білків. Селеноцистеїн може використовуватись в комбінації із селенометіоніном (SeMet). Селеноцистеїн із стабільний ізотоп 77Se (ядерний спін 1/2) застосовується в спектроскопії ЯРК.[2]
Див. також
- Пірролізин — 22-га протеїногенна амінокислота.
- Селенометіонін — інша селенвмісна амінокислота, яка випадковим чином вбудовується на місце метіоніну при біосинтезі білків.
Посилання
- IUPAC-IUBMB Joint Commission on Biochemical Nomenclature (JCBN) and Nomenclature Committee of IUBMB (NC-IUBMB) (PDF). European Journal of Biochemistry 264 (2). 1999. с. 607–609. doi:10.1046/j.1432-1327.1999.news99.x.
- Johansson, L.; Gafvelin, G.; Amér, E. S. J. (2005). Selenocysteine in Proteins — Properties and Biotechnological Use. Biochimica et Biophysica Acta 1726 (1). с. 1–13. doi:10.1016/j.bbagen.2005.05.010.
- Stadtman, Therese (8 березня 1974). Selenium Biochemistry. Science 183 (4128). с. 915–22. Bibcode:1974Sci...183..915S. PMID 4605100. doi:10.1126/science.183.4128.915.
- Roy, G.; Sarma, B. K.; Phadnis, P.P.; Mugesh, G. (2005). Selenium-containing enzymes in mammals: chemical perspectives. Journal of Chemical Sciences 117 (4). с. 287–303. doi:10.1007/BF02708441.
- Byun, B. J.; Kang, Y. K. (2011). Conformational Preferences and pKa Value of Selenocysteine Residue. Biopolymers 95 (5). с. 345–353. PMID 21213257. doi:10.1002/bip.21581.
- Longtin, R (2004). A forgotten debate: Is selenocysteine the 21st amino acid?. Journal of the National Cancer Institute 96 (7). с. 504–5. PMID 15069108. doi:10.1093/jnci/96.7.504.
- Böck A.; Forchhammer, K.; Heider, J.; Baron, C. (1991). Selenoprotein Synthesis: An Expansion of the Genetic Code. Trends in Biochemical Sciences 16 (12). с. 463–467. PMID 1838215. doi:10.1016/0968-0004(91)90180-4.
- Baranov P. V.; Gesteland R. F.; Atkins, J. F. (2002). Recoding: Translational Bifurcations in Gene Expression. Gene 286 (5). с. 187–201. PMID 11943474. doi:10.1016/S0378-1119(02)00423-7.
- Donovan, J.; Copeland, P. R. (2010). The Efficiency of Selenocysteine Incorporation is Regulated by Translation Initiation Factors. Journal of Molecular Biology 400 (4). с. 659–664. PMC 3721751. PMID 20488192. doi:10.1016/j.jmb.2010.05.026.
- Atkins, J. F. (2009). Recoding: Expansion of Decoding Rules Enriches Gene Expression. Springer. с. 31. ISBN 978-0-387-89381-5.
- Berry, M. J.; Banu, L.; Harney, J. W.; Larsen, P. R. (1993). Functional Characterization of the Eukaryotic SECIS Elements which Direct Selenocysteine Insertion at UGA Codons (PDF). The EMBO Journal 12 (8). с. 3315–3322. PMC 413599. PMID 8344267.
- Xu, Xue-Ming; Carlson, Bradley A; Mix, Heiko; Zhang, Yan; Saira, Kazima; Glass, Richard S; Berry, Marla J; Gladyshev, Vadim N та ін. Biosynthesis of Selenocysteine on Its tRNA in Eukaryotes. PLoS Biology 5 (1). PMC 1717018. PMID 17194211. doi:10.1371/journal.pbio.0050004.
- Yuan, Jing; Palioura, Sotiria; Salazar, Juan Carlos; Su, Dan; O'Donoghue, Patrick; Hohn, Michael J.; Cardoso, Alexander Machado; Whitman, William B. та ін. (12 грудня 2006). RNA-dependent conversion of phosphoserine forms selenocysteine in eukaryotes and archaea. Proceedings of the National Academy of Sciences (англ.) 103 (50). с. 18923–18927. ISSN 0027-8424. PMC 1748153. PMID 17142313. doi:10.1073/pnas.0609703104.
- Kryukov, G. V.; Castellano, S.; Novoselov, S. V.; Lobanov, A. V.; Zehtab, O.; Guigó, R.; Gladyshev, V. N. (2003). Characterization of Mammalian Selenoproteomes. Science 300 (5624). с. 1439–1443. Bibcode:2003Sci...300.1439K. PMID 12775843. doi:10.1126/science.1083516.
- Block, E. (2010). Garlic and Other Alliums: The Lore and the Science. Royal Society of Chemistry. ISBN 0-85404-190-7.
