Плазматична клітина
Плазматичні клітини або плазматичні В-клітини — лейкоцити, які виникають у лімфоїдних органах з В-лімфоцитів[1][2] і виділяють велику кількість білків, які називаються антитілами, у відповідь на специфічні речовини, які називаються антигенами. Ці антитіла переносяться з плазматичних клітин плазмою крові та лімфатичною системою до місця дії антигену-мішені (чужорідної речовини), де викликають його нейтралізацію або руйнування. В-клітини диференціюються в плазматичні клітини, які виробляють антитіла, схожі на рецептори В-клітини-попередника.[3]
| Плазматична клітина | |
|---|---|
| Деталі | |
| система | Лімфатична система |
| Ідентифікатори | |
| латинська | plasmacytos |
| MeSH | D010950 |
| TH | H2.00.03.0.01006 |
| FMA | 70574 |
Структура
Плазматичні клітини — це великі лімфоцити з великою кількістю базофільної цитоплазми і характерним виглядом під час світлової мікроскопії. Вони мають ексцентричне ядро з гетерохроматином у характерному розташуванні колеса або циферблата. Їх цитоплазма також містить бліду зону, яка на електронній мікроскопії містить розгалужений апарат Гольджі та центріолі (ЕМ-фото). Розгалужений ендоплазматичний ретикулум поєднанні з добре розвиненим апаратом Гольджі робить плазматичні клітини добре пристосованими до секреції імуноглобулінів.[4] Інші органели плазматичної клітини включають рибосоми, лізосоми, мітохондрії та плазматичну мембрану.
Поверхневі антигени
Остаточно диференційовані плазматичні клітини експресують відносно мало поверхневих антигенів і не експресують загальні маркери усіх В-клітин, такі як CD19 і CD20. Натомість плазматичні клітини ідентифікуються за допомогою проточної цитометрії за їх додатковою експресією CD138, CD78 та рецептора інтерлейкіну-6 . У людини CD27 є хорошим маркером для плазматичних клітин; наївні В-клітини — це CD27-, В-клітини пам'яті — це CD27+, а плазматичні клітини — CD27++.[5]
Поверхневий антиген CD138 (синдекан-1) експресується у високих рівнях.[6]
Іншим важливим поверхневим антигеном є CD319 (SLAMF7). Цей антиген експресується у високих рівнях на нормальних людських плазматичних клітинах. Він також виражений на злоякісних плазматичних клітинах при множинній мієломі. У порівнянні з CD138, який швидко зникає ex vivo, експресія CD319 значно стабільніша.[7]
Розвиток
Після виходу з кісткового мозку В-клітина діє як антигенпрезентувальна клітина (АПК) і поглинає (інтерналізує) антигени шляхом рецептор-опосередкованого ендоцитозу та обробляються (процесінг). Частинки антигену (які зараз відомі як антигенні пептиди) прикріпляються на молекули головного комплексу гістосумісності класу II (MHC II) і «представляються» на зовнішньоклітинній поверхні CD4+ Т-клітинам (іноді їх називають Т-хелперами). Ці Т-клітини зв'язуються з комплексом антиген-MHC II і викликають активацію В-клітини. Такий тип захисту системи, подібний до двофакторного методу аутентифікації. Спочатку В-клітини повинні зіткнутися з чужорідним антигеном, а потім вони повинні бути активовані Т-хелперами, перш ніж вони диференціюються в специфічні клітини.
Після стимуляції Т-клітиною, яка зазвичай відбувається в зародкових центрах вторинних лімфоїдних органів, таких як селезінка та лімфатичні вузли, активована В-клітина починає диференціюватися в більш спеціалізовані клітини. В-клітини зародкового центру можуть диференціюватися на В-клітини пам'яті або плазматичні клітини. Більшість із цих В-клітин стануть плазмабластами (або «незрілими» плазматичними клітинами) і, зрештою, плазматичними клітинами, і починають виробляти велику кількість антитіл. Деякі В-клітини зазнають процесу, відомого як «дозрівання спорідненості».[8] Цей процес сприяє активації та росту клонів В-клітин, здатних виділяти антитіла більш високої спорідненості до антигену, шляхом відбору на здатність зв'язувати антиген із вищою афінністю.
Незрілі плазматичні клітини
Найнезрілішою плазматичною клітиною, є плазмобласт.[9] Плазмобласти виділяють більше антитіл, ніж В-клітини, але менше, ніж плазматичні клітини.[10] Вони швидко діляться і все ще здатні інтерналізувати антигени та презентувати їх Т-клітинам.[10] Клітина може перебувати в цьому стані кілька днів, а потім або гине, або безповоротно диференціюється в зрілу, повністю диференційовану плазматичну клітину.[10] Диференціація зрілих В-клітин у плазматичні клітини залежить від факторів транскрипції Blimp-1/PRDM1 та IRF4 .
Функція
На відміну від своїх попередників, плазматичні клітини не можуть перемикати класи імуноглобулінів, не можуть діяти як антигенопрезентувальні клітини, оскільки вони більше не мають MHCII на своїй поверхні, і не поглинають антиген, оскільки вони більше не мають значну кількість імуноглобуліну на своїй поверхні.[10] Проте постійний вплив антигену через навіть ці низькі рівні імуноглобуліну на поверхні є важливим, оскільки це частково визначає тривалість життя клітини.[10]
Тривалість життя, клас вироблених антитіл і місце, куди рухається плазматична клітина, також залежать від сигналів, таких як цитокіни, отримані від Т-клітини під час диференціювання.[11] Диференціація за допомогою Т-клітино-незалежної антигенної стимуляції (стимуляція В-клітини, яка не вимагає участі Т-клітини) може відбуватися в будь-якому місці тіла[8] і призводить до появи короткоживучих клітин, які виділяють антитіла IgM.[11] Процеси, залежні від Т-клітин, поділяються на первинні та вторинні реакції: первинна відповідь (це означає, що Т-клітина присутня в момент первинного контакту В-клітини з антигеном) приводить до появи короткоживучих клітин, які залишаються в екстрамедулярних областях лімфатичних вузлів; вторинна реакція призводить до появи клітин, які живуть триваліше, виробляють IgG та IgA і часто подорожують до кісткового мозку.[11] Наприклад, плазматичні клітини, ймовірно, будуть секретувати антитіла IgG3, якщо вони дозрівають у присутності цитокінів-інтерферону-гамма. Оскільки дозрівання В-клітин також включає соматичну гіпермутацію (процес, який завершується до диференціювання в плазматичні клітини), ці антитіла часто мають дуже високу спорідненість до свого антигену.
Плазматичні клітини можуть виробляти лише один вид антитіл в одному класі імуноглобуліну. Іншими словами, кожна В-клітина специфічна для одного антигену, але кожна клітина може виробляти кілька тисяч відповідних антитіл за секунду.[12] Це рясне вироблення антитіл є невід'ємною частиною гуморальної імунної відповіді.
Довгоживучі плазматичні клітини
Результати сучасних досліджень свідчать про те, що після процесу дозрівання спорідненості в зародкових центрах плазматичні клітини розвиваються в один із двох типів клітин: короткоживучі плазматичні клітини (short-lived plasma cells, SLPC) або довгоживучі плазматичні клітини (long-lived plasma cells, LLPC). LLPC в основному знаходяться в кістковому мозку протягом тривалого періоду часу і виділяють антитіла, забезпечуючи таким чином довготривалий захист. LLPC може підтримувати вироблення антитіл протягом десятиліть або навіть протягом усього життя людини,[13][14] і, на відміну від В-клітин, LLPC не потребують повторної стимуляції антигеном для вироблення антитіл. Людську популяцію LLPC можна ідентифікувати як клітини CD19 – CD38hi CD138+.[15]
Довгострокове виживання LLPC залежить від конкретного середовища (ніші) в кістковому мозку.[16] Видалення LLPC з їх ніші призводить до їх швидкої смерті. Ніша виживання може підтримувати лише обмежену кількість LLPC, тому середовище ніші має захищати свої клітини LLPC, але все ж надавати можливість адаптуватися до нових надходжень.[17][18] Ніша виживання плазматичних клітин визначається комбінацією клітинних і молекулярних факторів, і хоча її ще не визначено належним чином, було ідентифіковано кілька молекул, які підтримують виживання LLPC, таких як IL-5, IL-6, TNF-α, фактор-1α, отриманий із стромальних клітин, і передача сигналів через CD44.[19] LLPC також в меншій мірі можна знайти в лімфоїдній тканині, асоційованій з кишечником (GALT), де вони виробляють антитіла IgA і сприяють імунітету слизової оболонки. Останні результати свідчать про те, що плазматичні клітини в кишечнику не обов'язково повинні бути створені de novo з активних В-клітин, й існують також LLPC, що вимагає таку ж нішу виживання, як і в кістковому мозку.[20] Тканиноспецифічні ніші, які дозволяють виживати LLPC, також були описані в назально-асоційованих лімфоїдних тканинах (NALT), тонзилярних лімфоїдних тканинах людини та слизовій оболонці людини або лімфоїдних тканинах, асоційованих зі слизовою оболонкою (MALT)[21][22][23][24]
Спочатку вважалося, що безперервне вироблення антитіл є результатом постійного поповнення SLPC шляхом повторної стимуляції В-клітин пам'яті. Однак останні дослідження показують, що деякі плазматичні клітини дійсно довговічні. Дослідження показали, що відсутність антигену та виснаження В-клітин не впливають на вироблення високоафінних антитіл LLPC. Тривале виснаження В-клітин (при лікуванні моноклональними антитілами проти CD20, яке діє на В-клітини, але не плазматичні клітини) також не вплинуло на титри антитіл.[25][26][27] LLPC секретує високі рівні IgG незалежно від В-клітин. LLPC в кістковому мозку є основним джерелом циркулюючих IgG у людини.[28] Незважаючи на те, що виробництво IgA традиційно пов'язане зі слизовими оболонками, деякі плазматичні клітини в кістковому мозку також виробляють IgA.[29] Ми також можемо знайти LLPC в кістковому мозку, які виробляють IgM.[30]
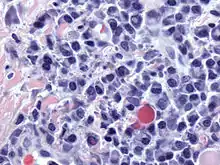
Клінічне значення
Плазмоцитома, мієломна хвороба, макроглобулінемія Вальденстрема, хвороба важких ланцюгів і лейкемія плазматичних клітин є злоякісними новоутвореннями плазматичних клітин.[31] Мієломну хворобу виявляють часто, оскільки злоякісні плазматичні клітини продовжують виробляти антитіло, яке можна виявити як парапротеїн .
Вважається, що звичайний варіабельний імунодефіцит пов'язаний з проблемою диференціювання лімфоцитів у плазматичні клітини. Результатом є низький рівень антитіл у сироватці крові та ризик інфекцій.
Первинний амілоїдоз спричиняється відкладенням надлишку легких ланцюгів імуноглобуліну, які виділяються з плазматичних клітин.
Див. також
- Дискразія плазматичних клітин
- Лейкоцити
Посилання
- Guyton and Hall Textbook of Medical Physiology 14th edition: unit 6, chapter 35.
- Plasma Cell - an overview | ScienceDirect Topics.
- Plasma cell - biology. britannica.com.
- Plasma Cell - LabCE.com, Laboratory Continuing Education. www.labce.com. Процитовано 2 червня 2018.
- 5. Textbook of Immunology (вид. 2). CRC Press. 1996. с. 102. ISBN 978-3-7186-0596-5.
- Rawstron AC (May 2006). Immunophenotyping of plasma cells. Current Protocols in Cytometry. Chapter. Chapter 6: Unit6.23. ISBN 0-471-14295-6. PMID 18770841. doi:10.1002/0471142956.cy0623s36.
- Frigyesi I, Adolfsson J, Ali M, Christophersen MK, Johnsson E, Turesson I, Gullberg U, Hansson M, Nilsson B (February 2014). Robust isolation of malignant plasma cells in multiple myeloma. Blood 123 (9): 1336–40. PMID 24385542. doi:10.1182/blood-2013-09-529800. Проігноровано невідомий параметр
|doi-access=(довідка) - Molecular biology of B cells. Amsterdam: Elsevier. 2004. с. 189–191. ISBN 0-12-053641-2.
- Wintrobe's Clinical Hematology, 2-Vol. Set. Hagerstwon, MD: Lippincott Williams & Wilkins. 2008. с. 347. ISBN 978-0-7817-6507-7.
- Janeway's immunobiology. New York: Garland Science. 2008. с. 387–388. ISBN 978-0-8153-4123-9.
- Human B Cell Populations (Chemical Immunology) 67. S. Karger AG (Switzerland). 1997. с. 103–104. ISBN 3-8055-6460-0.
- Histology and cell biology: an introduction to pathology. St. Louis: Mosby. 2002. с. 275. ISBN 0-323-01639-1.
- Slifka MK, Matloubian M, Ahmed R (March 1995). Bone marrow is a major site of long-term antibody production after acute viral infection. Journal of Virology 69 (3): 1895–902. PMC 188803. PMID 7853531. doi:10.1128/jvi.69.3.1895-1902.1995.
- Radbruch, Andreas; Muehlinghaus, Gwendolin; Luger, Elke O.; Inamine, Ayako; Smith, Kenneth G. C.; Dörner, Thomas; Hiepe, Falk (October 2006). Competence and competition: the challenge of becoming a long-lived plasma cell. Nature Reviews Immunology (англ.) 6 (10): 741–750. ISSN 1474-1733. PMID 16977339. doi:10.1038/nri1886.
- Halliley JL, Tipton CM, Liesveld J, Rosenberg AF, Darce J, Gregoretti IV, Popova L, Kaminiski D, Fucile CF, Albizua I, Kyu S, Chiang KY, Bradley KT, Burack R, Slifka M, Hammarlund E, Wu H, Zhao L, Walsh EE, Falsey AR, Randall TD, Cheung WC, Sanz I, Lee FE (July 2015). Long-Lived Plasma Cells Are Contained within the CD19(-)CD38(hi)CD138(+) Subset in Human Bone Marrow. Immunity 43 (1): 132–45. PMC 4680845. PMID 26187412. doi:10.1016/j.immuni.2015.06.016.
- Manz RA, Radbruch A (April 2002). Plasma cells for a lifetime?. European Journal of Immunology 32 (4): 923–7. PMID 11920557. doi:10.1002/1521-4141(200204)32:4<923::aid-immu923>3.0.co;2-1. Проігноровано невідомий параметр
|doi-access=(довідка) - Nguyen, Doan C.; Joyner, Chester J.; Sanz, Iñaki; Lee, F. Eun-Hyung (11 вересня 2019). Factors Affecting Early Antibody Secreting Cell Maturation Into Long-Lived Plasma Cells. Frontiers in Immunology 10: 2138. ISSN 1664-3224. PMC 6749102. PMID 31572364. doi:10.3389/fimmu.2019.02138. Проігноровано невідомий параметр
|doi-access=(довідка) - Tangye, Stuart G. (December 2011). Staying alive: regulation of plasma cell survival. Trends in Immunology (англ.) 32 (12): 595–602. PMID 22001488. doi:10.1016/j.it.2011.09.001.
- Cassese G, Arce S, Hauser AE, Lehnert K, Moewes B, Mostarac M, Muehlinghaus G, Szyska M, Radbruch A, Manz RA (August 2003). Plasma cell survival is mediated by synergistic effects of cytokines and adhesion-dependent signals. Journal of Immunology 171 (4): 1684–90. PMID 12902466. doi:10.4049/jimmunol.171.4.1684. Проігноровано невідомий параметр
|doi-access=(довідка) - Lemke A, Kraft M, Roth K, Riedel R, Lammerding D, Hauser AE (January 2016). Long-lived plasma cells are generated in mucosal immune responses and contribute to the bone marrow plasma cell pool in mice. Mucosal Immunology 9 (1): 83–97. PMID 25943272. doi:10.1038/mi.2015.38. Проігноровано невідомий параметр
|doi-access=(довідка) - Liang, Bin; Hyland, Lisa; Hou, Sam (June 2001). Nasal-Associated Lymphoid Tissue Is a Site of Long-Term Virus-Specific Antibody Production following Respiratory Virus Infection of Mice. Journal of Virology (англ.) 75 (11): 5416–5420. ISSN 0022-538X. PMC 114951. PMID 11333927. doi:10.1128/JVI.75.11.5416-5420.2001.
- van Laar, Jacob M.; Melchers, Marc; Teng, Y. K. Onno; van der Zouwen, Boris; Mohammadi, Rozbeh; Fischer, Randy; Margolis, Leonid; Fitzgerald, Wendy та ін. (September 2007). Sustained Secretion of Immunoglobulin by Long-Lived Human Tonsil Plasma Cells. The American Journal of Pathology (англ.) 171 (3): 917–927. PMC 1959503. PMID 17690187. doi:10.2353/ajpath.2007.070005.
- Huard, Bertrand; McKee, Thomas; Bosshard, Carine; Durual, Stéphane; Matthes, Thomas; Myit, Samir; Donze, Olivier; Frossard, Christophe та ін. (1 липня 2008). APRIL secreted by neutrophils binds to heparan sulfate proteoglycans to create plasma cell niches in human mucosa. Journal of Clinical Investigation (англ.) 118 (8): 2887–2895. ISSN 0021-9738. PMC 2447926. PMID 18618015. doi:10.1172/JCI33760.
- Lemke, A; Kraft, M; Roth, K; Riedel, R; Lammerding, D; Hauser, A E (January 2016). Long-lived plasma cells are generated in mucosal immune responses and contribute to the bone marrow plasma cell pool in mice. Mucosal Immunology (англ.) 9 (1): 83–97. ISSN 1933-0219. PMID 25943272. doi:10.1038/mi.2015.38.
- Slifka MK, Antia R, Whitmire JK, Ahmed R (March 1998). Humoral immunity due to long-lived plasma cells. Immunity 8 (3): 363–72. PMID 9529153. doi:10.1016/S1074-7613(00)80541-5. Проігноровано невідомий параметр
|doi-access=(довідка) - DiLillo DJ, Hamaguchi Y, Ueda Y, Yang K, Uchida J, Haas KM, Kelsoe G, Tedder TF (January 2008). Maintenance of long-lived plasma cells and serological memory despite mature and memory B cell depletion during CD20 immunotherapy in mice. Journal of Immunology 180 (1): 361–71. PMID 18097037. doi:10.4049/jimmunol.180.1.361. Проігноровано невідомий параметр
|doi-access=(довідка) - Ahuja A, Anderson SM, Khalil A, Shlomchik MJ (March 2008). Maintenance of the plasma cell pool is independent of memory B cells. Proceedings of the National Academy of Sciences of the United States of America 105 (12): 4802–7. Bibcode:2008PNAS..105.4802A. PMC 2290811. PMID 18339801. doi:10.1073/pnas.0800555105. Проігноровано невідомий параметр
|doi-access=(довідка) - Longmire RL, McMillan R, Yelenosky R, Armstrong S, Lang JE, Craddock CG (October 1973). In vitro splenic IgG synthesis in Hodgkin's disease. The New England Journal of Medicine 289 (15): 763–7. PMID 4542304. doi:10.1056/nejm197310112891501.
- Mei HE, Yoshida T, Sime W, Hiepe F, Thiele K, Manz RA, Radbruch A, Dörner T (March 2009). Blood-borne human plasma cells in steady state are derived from mucosal immune responses. Blood 113 (11): 2461–9. PMID 18987362. doi:10.1182/blood-2008-04-153544.
- Bohannon C, Powers R, Satyabhama L, Cui A, Tipton C, Michaeli M, Skountzou I, Mittler RS, Kleinstein SH, Mehr R, Lee FE, Sanz I, Jacob J (June 2016). Long-lived antigen-induced IgM plasma cells demonstrate somatic mutations and contribute to long-term protection. Nature Communications 7 (1): 11826. Bibcode:2016NatCo...711826B. PMC 4899631. PMID 27270306. doi:10.1038/ncomms11826.
- "Plasma cell" на вебсайті Dorland's Medical Dictionary

{kind=link}