Поясна кора
Поясна кора — глибинна ділянка, розташована в медіальній частині кори головного мозку. Поясна кора складається з поясної звивини (лат. gyrus cinguli), яка лежить безпосередньо над мозолистим тілом, і поясної борозни (лат. sulcus cinguli). Поясна кора зазвичай вважається частиною обідкової або лімбічної частки (яку останнім часом в літературі виділяють окремо, ставлячи в один ряд з лобовою, тім'яною, скроневою й потиличною).
| Поясна кора | |
|---|---|
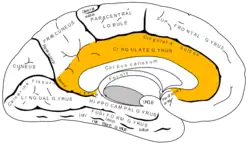 Медіальна поверхня лівої півкулі головного мозку з ввиділеним зображенням поясної звивини. | |
| Деталі | |
| Частина від | Cerebral cortex |
| Артерія | Передня мозкова артерія |
| Вена | Верхній сагітальний сінус. |
| Ідентифікатори | |
| Латина | Cortex cingularis, gyrus cinguli |
| Акронім(и) | Cg |
| Анатомія Грея | subject #189 |
| MeSH | D006179 |
| NeuroNames | 159 |
| NeuroLex ID | birnlex_798 |
| TA98 | A14.1.09.231 |
| TA2 | 5513 |
| FMA | 62434[1] |
| Анатомічна термінологія | |
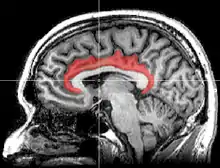
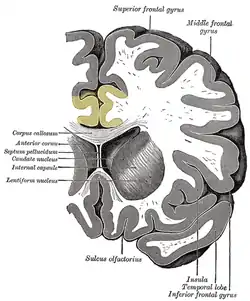
Поясна кора отримує вхідні імпульси від таламуса та кори головного мозку[2], а також, у свою чергу, надсилає вихідні імпульси до енторинальної кори через шляхи білої речовини, які мають назву поясу (лат. cingulum). Поясна кора — невід'ємна частина лімбічної системи, яка бере участь у формуванні й обробці емоцій[3] навчання[4] і пам'яті[5][6]. Поєднання цих трьох функцій робить поясну звивину досить значущою у прив'язці поведінкових результатів до мотивації (наприклад, певна дія зумовлює позитивний емоційний відгук, в результаті навчання)[7]. Ця роль робить поясну кору дуже важливою в патогенезі таких розладів, як депресія[8] та шизофренія. Вона також відіграє роль у виконавчій функції і дихальному контролі[9].
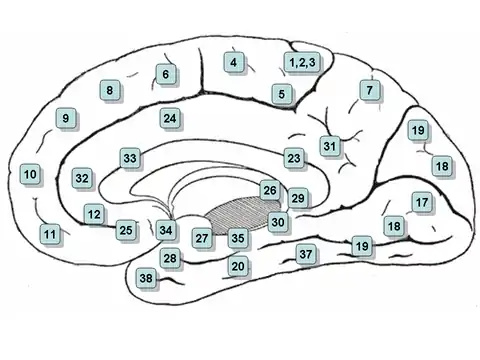
Структура
На основі цитоархітектоніки поясна кора поділяється на поля Бродмана 23, 24, 26, 29, 30, 31, 32 і 33. Поля 26, 29 та 30 зазвичай ще називають ретроспленальними.
Передня поясна кора
Ця назва відповідає полям 24, 32 і 33 Бродмана (а також Костянтина фон Економо, Бейлі і фон Боніна).
Вона протягається допереду до поля Бродмана 25 (розташованого нижче за коліно мозолистого тіла). Передня поясна кора бере участь у виявленні конфліктних і помилкових процесів.[10]
Задня поясна кора
Відповідає полям 23 і 31 Бродмана . Її клітинна структура зерниста. Її продовженням дозаду є ретроспленальна кора (поле Бродмана 29); дорзально — гранулярне поле Бродмана 31. Задня частина поясної звивини отримує більшу частину своїх аферентних аксонів від поверхневих ядер таламуса, який сам отримує аксони від субікулярної ділянки (лат. subiculum). Зниження обміну речовин (гіпометаболізм) задньої частини поясної кори (з 18F-FDG PET) було помічено при хворобі Альцгеймера.
Вхідні імпульси до передньої поясної звивини
Експерименти на макаках показали, що вентральні передня й вентральні латеральні ядра таламуса пов'язані з моторними ділянками поясної борозни.[11][12].
Вихідні імпульси від передньої поясної звивини
Ростральна частина поясної звивини (поле Бродмана 32) шле імпульси до ростральної частини верхньої скроневої звивини, міжорбітальної кори і латеральної префронтальної кори.[13]
Вентральна передня частина поясної звивини (поле Бродмана 24) відправляє імпульси до передньої острівцевої кори, премоторної кори (поле Бродмана 6), поля Бродмана 8, навколонюхову ділянку, очноямкову кіркову ділянку (поле Бродмана 12), В латеробазальні ядра мигдалеподібного тіла, і ростральну частину нижньої тім'яної часточки. Експерименти на котах показали, що передня поясна звивина має двосторонні зв'язки з ростральною частиною таламуса, заднім латеральним ядром і ростральним краєм подушки зорового бугра.[14] .
Вихідні імпульси від задньої поясної звивини
Задня частина поясної звивини кори головного мозку (поле Бродмана 23) посилає імпульси в дорсолатеральну префронтальну кору (поле Бродмана 9), передню префронтальну кору (поле Бродмана 10), орбітофронтальну кору (поле Бродмана 11), в приморськоконикову звивину (лат. gyrus parahyppocampalis), задню частину нижньої тім'яної часточки (лат. lobulus parietalis), в пресубікулярну ділянку (лат. presubiculum), верхню скроневу борозну і ретроспленальну ділянку.[13]
Вентральна задня частина поясної звивини кори, як встановлено, взаємно з'єднана з каудальною частиною задньої тім'яної частки у макак-резус.Також медіальна задня тім'яна кора пов'язана з задньою вентральною частиною поясної борозни.[15][16][17]
Клінічне значення
Шизофренія
Використовуючи тривимірну магнітно-резонансну томографію для вимірювання об'єму передньої ростральної частини поясної звивини (біля коліна мозолистого тіла), Такагаші та співавт. (2003) виявили, що її обсяг більше в здоровій контрольній групі у жінок, ніж у чоловіків, але не було виявлено такої статевої різниці у людей з шизофренією. Люди з шизофренією також мали загалом менший обсяг передньої ростральної частини поясної звивини, ніж здорові особи в контрольній групі.[18]
Haznedar і співавт. (2004) вивчали інтенсивність метаболізму глюкози в передній і задній частинах поясної звивини у пацієнтів з шизофренією, шизотиповим розладом особистості й порівнювали їх із контрольною групою. Швидкість метаболізму глюкози виявився нижче у пацієнтів з шизофренією в порівнянні з контрольною групою. Хоча метаболізм глюкози улюдей з шизотиповим розладом особистості очікувався десь посередині між хворими шизофренією і контрольною групою, вони насправді мали більш високий метаболізм глюкози рівень лівої задньої поясної звивини. Обсяг лівої передньої поясної звивини був знижений у пацієнтів з шизофренією в порівнянні з контролем, але не було ніякої різниці між людьми з шизотиповим розладом особистості та людей з шизофренією. З цих результатів випливає, що шизофренія і шизотиповий розлад — це дві різних нозологічних одиниці.[19]
Дослідження обсягу сірої і білої речовини в передньої поясної звивини у пацієнтів з шизофренією та їх здорових родичів першого та другого ступеня виявили суттєвої різниці в обсязі білого речовини у людей з шизофренією та їхніх здорових родичів. Як хворі на шизофренію так і їхні здорові родичі першого ступеня мають менше сірої речовини, ніж здорові родичі другого ступеня. Виявилося, що гени відповідають за зниження обсягу сірої речовини у людей з шизофренією.[20]
Фудзівара і співавт. (2007) провели експеримент, в якому вони співвіднесли розмір передньої частини поясної звивини у пацієнтів із шизофренією з їх функціонуванням у соціальному пізнанні, психопатології і емоції з контрольною групою. Чим менше Розмір передньої частини поясної звивини, тим нижчий рівень соціального функціонування і тим вище був психопатології у людей з шизофренією.[21]
Резюме
Люди з шизофренією мають відмінності передньої поясної звивини в порівнянні з контрольною групою. Передня поясна звивина виявилася менше у людей з шизофренією.[21] Обсяг сірої речовини в передній поясній звивині виявився нижче у осіб із шизофренією.[19][20] Здорові жінки мають більшу ростральну частину передньої поясної звивини, ніж чоловіки, але ця статева різниця в розмірі відсутня у людей з шизофренією.[18] Швидкість метаболізму глюкози нижче в лівій передній поясній звивині і в правій задній поясній звивині.[19]
На додаток до змін в передній поясній корі, інші структури головного мозку також показують зміни в випадках шизофренії в порівнянні з контрольною групою. Морський коник у пацієнтів з шизофренією виявився меншим за розміром порівняно з контрольною групою того ж віку,[22] і, аналогічно, хвостате ядро й бліда куля також виявилися меншими за обсягом в дослідженні людей з шизофренією.[23] У той час як у людей з шизофренією обсяг сірої речовини зменшений, розміри бічних і третього шлуночків збільшені.[24]
Додаткові зображення
 Медіальна поверхня півкулі головного мозку. Медіальний вигляд. Глибокий розтин.
Медіальна поверхня півкулі головного мозку. Медіальний вигляд. Глибокий розтин. Медіальна поверхня півкулі головного мозку. Медіальний вигляд. Глибокий розтин.
Медіальна поверхня півкулі головного мозку. Медіальний вигляд. Глибокий розтин. Медіальна поверхня півкулі головного мозку. Медіальний вигляд. Глибокий розтин.
Медіальна поверхня півкулі головного мозку. Медіальний вигляд. Глибокий розтин.
Посилання
- Foundational Model of Anatomy
- Oстаннім часом англомовна література з нейронауки (англ. neuroscience) рясніє термінологією з електронної схемотехніки щодо нервових структур і аферентні сигнали називають вхідними (англ. input) а еферентні — вихідними (англ. output).
- Hadland, K. A.; Rushworth M.F. (2003). The effect of cingulate lesions on social behaviour and emotion. Neuropsychologia 41 (8): 919–931. PMID 12667528. doi:10.1016/s0028-3932(02)00325-1.
- Cingulate binds learning. Trends Cogn Sci 1 (1): 2. 1997. PMID 21223838. doi:10.1016/s1364-6613(97)85002-4.
- Kozlovskiy, S.; Vartanov A.; Pyasik M.; Nikonova E.; Velichkovsky B. (10 жовтня 2013). Anatomical Characteristics of Cingulate Cortex and Neuropsychological Memory Tests Performance. Procedia - Social and Behavioral Sciences 86: 128–133. doi:10.1016/j.sbspro.2013.08.537.
- Kozlovskiy, S.A.; Vartanov A.V.; Nikonova E.Y.; Pyasik M.M.; Velichkovsky B.M. (2012). The Cingulate Cortex and Human Memory Processes. Psychology in Russia: State of the Art 5: 231–243. doi:10.11621/pir.2012.0014.
- Hayden, B. Y.; Platt, M. L. (2010). Neurons in Anterior Cingulate Cortex Multiplex Information about Reward and Action. Journal of Neuroscience 30 (9): 3339–3346. PMC 2847481. PMID 20203193. doi:10.1523/JNEUROSCI.4874-09.2010.
- Drevets, W. C.; Savitz, J.; Trimble, M. (2008). The subgenual anterior cingulate cortex in mood disorders. CNS spectrums 13 (8): 663–681. PMC 2729429. PMID 18704022. doi:10.1017/s1092852900013754.
- Adams, R.; David, A. S. (2007). Patterns of anterior cingulate activation in schizophrenia: A selective review. Neuropsychiatric disease and treatment 3 (1): 87–101. PMC 2654525. PMID 19300540. doi:10.2147/nedt.2007.3.1.87.
- Dorland's. Illustrated medical dictionary. Elsevier Saunders. с. 363. ISBN 978-14160-6257-8.
- McFarland, N. R.; Harber, S. N. (2000). Convergent Inputs from Thalamic Motor Nuclei and Frontal Cortical Areas to the Dorsal Striatum in the Primate. The Journal of Neuroscience 20 (10): 3798–3813.
- Wyass, J. M.; Van Groen, T. (1992). Connections between the retrosplenial cortex and the hippocampal formation in the rat: A review. Hippocampus 2 (1): 1–11. PMID 1308170. doi:10.1002/hipo.450020102.
- Pandya, D. N.; Hoesen, G. W.; Mesulam, M. -M. (1981). Efferent connections of the cingulate gyrus in the rhesus monkey. Experimental Brain Research. 42–42 (3–4): 319–330. doi:10.1007/BF00237497.
- Fujii, M. (1983). Fiber connections between the thalamic posterior lateral nucleus and the cingulate gyrus in the cat. Neuroscience Letters 39 (2): 137–142. PMID 6688863. doi:10.1016/0304-3940(83)90066-6.
- Petrides, M; Pandya, DN (24 жовтня 2007). Efferent association pathways from the rostral prefrontal cortex in the macaque monkey.. The Journal of neuroscience : the official journal of the Society for Neuroscience 27 (43): 11573–86. PMID 17959800. doi:10.1523/JNEUROSCI.2419-07.2007.
- Cavada, C.; Goldman-Rakic, P. S. (1989). Posterior parietal cortex in rhesus monkey: I. Parcellation of areas based on distinctive limbic and sensory corticocortical connections. The Journal of Comparative Neurology 287 (4): 393–421. PMID 2477405. doi:10.1002/cne.902870402.
- Adey, W. R. (1951). An Experimental Study of the Hippocampal Connexions of the Cingulate Cortex in the Rabbit. Brain 74 (2): 233–247. PMID 14858747. doi:10.1093/brain/74.2.233.
- Takahashi, T.; Suzuki, M.; Kawasaki, Y.; Hagino, H.; Yamashita, I.; Nohara, S.; Nakamura, K.; Seto, H. та ін. (2003). Perigenual cingulate gyrus volume in patients with schizophrenia: A magnetic resonance imaging study. Biological Psychiatry 53 (7): 593–600. PMID 12679237. doi:10.1016/S0006-3223(02)01483-X.
- Haznedar, M. M.; Buchsbaum, M. S.; Hazlett, E. A.; Shihabuddin, L.; New, A.; Siever, L. J. (2004). Cingulate gyrus volume and metabolism in the schizophrenia spectrum. Schizophrenia Research 71 (2–3): 249–262. PMID 15474896. doi:10.1016/j.schres.2004.02.025.
- Costain, G.; Ho, A.; Crawley, A. P.; Mikulis, D. J.; Brzustowicz, L. M.; Chow, E. W. C.; Bassett, A. S. (2010). Reduced gray matter in the anterior cingulate gyrus in familial schizophrenia: A preliminary report. Schizophrenia Research 122 (1–3): 81–84. PMC 3129334. PMID 20638248. doi:10.1016/j.schres.2010.06.014.
- Fujiwara, H.; Hirao, K.; Namiki, C.; Yamada, M.; Shimizu, M.; Fukuyama, H.; Hayashi, T.; Murai, T. (2007). Anterior cingulate pathology and social cognition in schizophrenia: A study of gray matter, white matter and sulcal morphometry. NeuroImage 36 (4): 1236–1245. PMID 17524666. doi:10.1016/j.neuroimage.2007.03.068.
- {Cite journal | last1 = Koolschijn | first1 = P. C. D. M. P. | last2 = Van Haren | first2 = N. E. M. | last3 = Cahn | first3 = W. | last4 = Schnack | first4 = H. G. | last5 = Janssen | first5 = J. | last6 = Klumpers | first6 = F. | last7 = Hulshoff Pol | first7 = H. E. | last8 = Kahn | first8 = R. S. | doi = 10.4088/JCP.08m04574yel | title = Hippocampal Volume Change in Schizophrenia | journal = The Journal of Clinical Psychiatry | volume = 71 | issue = 6 | pages = 737—744 | year = 2010 | pmid = 20492835| pmc = }}
- Mitelman, S. A.; Canfield, E. L.; Chu, K. W.; Brickman, A. M.; Shihabuddin, L.; Hazlett, E. A.; Buchsbaum, M. S. (2009). Poor outcome in chronic schizophrenia is associated with progressive loss of volume of the putamen. Schizophrenia Research 113 (2–3): 241–245. PMC 2763420. PMID 19616411. doi:10.1016/j.schres.2009.06.022.
- Kempton, M. J.; Stahl, D.; Williams, S. C. R.; Delisi, L. E. (2010). Progressive lateral ventricular enlargement in schizophrenia: A meta-analysis of longitudinal MRI studies. Schizophrenia Research 120 (1–3): 54–62. PMID 20537866. doi:10.1016/j.schres.2010.03.036.