Хромосомні аберації
Хромосо́мні абера́ції (абера́ції хромосо́много ти́пу, хромосомні мутації, хромосомні перебудови) — порушення структури хромосом, які відбуваються синхронно в обох хроматидах. Класифікують делеції (видалення ділянки хромосоми), інверсії (зміна порядку генів ділянки хромосоми на зворотний), дуплікації (повторення ділянки хромосоми), транслокації (перенесення ділянки хромосоми на іншу). Хромосомні перебудови носять, як правило, патологічний характер і нерідко призводять до загибелі організму. Показано значення хромосомних перебудов у видоутворенні та еволюції.
Виникнення хромосомних аберацій
У ході кросинговеру утворюються розриви хромосом, які потім репаруються. Порушення процесу репарації можуть призвести до появи хромосомних перебудов. Розриви хромосом і, як наслідок, утворення перебудов відбуваються під дією різних мутагенних факторів: фізичної (іонізуюче випромінювання), хімічної або біологічної (транспозони, віруси) природи. Також деякі хромосомні перебудови (аберації) характерні для носіїв специфічних сайтів ламкості.
Делеції
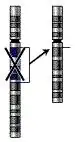
Розрізняють термінальні (втрата кінцевої ділянки хромосоми) і інтеркалярні (втрата частини хромосоми у її внутрішній ділянці) делеції. Якщо після утворення делеції хромосома зберегла центромеру, вона аналогічно до інших хромосом передається при поділі, ділянки без центромери як правило втрачаються. При кон'югації гомологів під час кросинговеру у нормальної хромосоми на місці делеції в мутантній хромосомі утворюється так звана делеційна петля, яка компенсує відсутність делетованої ділянки.
Досліджені делеції рідко захоплюють протяжні ділянки хромосом, зазвичай такі аберації летальні. Найкраще вивченим захворюванням, зумовленим делецією, є синдром котячого крику, описаний в 1963 році Джеромом Леженом. У його основі лежить делеція невеликої ділянки короткого плеча 5 хромосоми. Для хворих характерний ряд відхилень від норми: порушення функцій серцево-судинної, травної систем, недорозвинення гортані (з характерним криком, що нагадує котяче нявкання), загальне відставання розвитку, розумова відсталість, місяцеподібне обличчя з широко розставленими очима. Синдром зустрічається в 1 з 50000 новонароджених.
Іншим цікавим прикладом цього виду хромосомних аберацій є делеція в гені, що кодує рецептор CCR5. Цей рецептор використовується вірусом імунодефіциту людини (ВІЛ) для розпізнавання своїх Т-лімфоцитів. Продукт гену з делецією отримав назву CCR5-Δ32, цей варіант CCR5 не розпізнається ВІЛ, і носії такої мутації до ВІЛ несприйнятливі (це близько 10% європейців).
Дуплікації
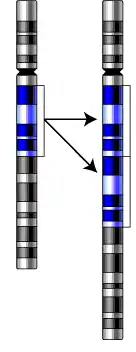
Дуплікації з'являються в результаті нерівного кросинговеру (в цьому випадку другий гомолог несе делецію) або в результаті помилки в ході реплікації. При кон'югації хромосоми з дуплікацією і нормальної хромосоми як і при делеції формується компенсаційна петля.
Практично у всіх організмів в нормі спостерігається множинність генів, що кодують рРНК (рибосомальні РНК). Це явище назвали надлишковістю генів. Так у E. coli на рДНК (ДНК, що кодує рРНК) припадає 0,4 % всього геному, що відповідає 5-10 копіям рибосомальних генів.
Інший приклад дуплікації — мутація Bar у Drosophila , виявлена у 20-х роках XX століття Т. Морганом та А. Стертевантом. Мутація зумовлена дуплікацією локусу 57.0 X-хромосоми. У нормальних самиць (B+/B+) очі мають по 800 фасеток, у гетерозиготних самиць (B+/B) очі мають по 350 фасеток, у гомозигот по мутації (B/B) — всього 70 фасеток. Виявлені також самиці з тричі повтореним ге́ном — double Bar (BD/B+).
У 1970 році Сусуму Оно в монографії «Еволюція шляхом дуплікації генів» розробив гіпотезу про еволюційну роль дуплікацій, які постачають нові гени, не зачіпаючи при цьому функцій вихідних генів. На користь цієї ідеї говорить близькість ряду генів по нуклеотидному складу, що кодують різні продукти. Це трипсин та хімотрипсин, гемоглобін та міоглобін та ряд інших білків.
Інверсії
Розрізняють парацентричні (інвертований фрагмент лежить по один бік від центромери) і перицентричні (інвертований фрагмент лежить по різні сторони від центромери) інверсії. При інверсії не відбувається втрати генетичного матеріалу, тому як такі інверсії як правило не впливають на фенотип, але якщо в інверсійної гетерозиготи (тобто організмі, що несе як нормальну хромосому, так і хромосому з інверсією) відбувається кросинговер, то існує ймовірність формування аномальних хроматид. У разі парацентричної інверсії утворюється одна нормальна і одна інвертована (фенотипово нормальна) хроматиди, дицентрична хроматида з дуплікацією і делецією (при розходженні хроматид вона зазвичай розривається на дві) і ацентрична хроматида з дуплікацію і делецією (зазвичай втрачається). У разі перицентричної інверсії утворюється одна нормальна і одна інвертована хроматиди, а також дві хроматиди з дуплікацію і делеція. Гамети, що несуть дефектні хромосоми, звичайно не розвиваються або гинуть на ранніх етапах ембріогенезу. Але гамети з інвертованою хромосомою розвиваються в організми, 50% гамет яких нежиттєздатні. Таким чином мутація зберігається в популяції.
У людини найпоширенішою є інверсія в 9 хромосомі, що не шкодить носієві, хоча існують дані, що у жінок з цією мутацією існує 30 % імовірність викидню.
Транслокації
Крім переносів ділянок з однієї негомологічної хромосоми на іншу, класифікують також реципрокні транслокації (коли дві негомологічні хромосоми обмінюються ділянками), робертсонівські транслокації(при цьому дві негомологічні хромосоми об'єднуються в одну), а також транспозиції (перенесення ділянки хромосоми на інше місце на тій же хромосомі).
Транслокація, реципрокна транслокація і транспозиція, які не супроводжуються втратою генетичного матеріалу (збалансовані транслокації), часто не виявляються фенотипово. Однак, як і у випадку з інверсіями, в процесі гаметогенезу частина сформованих гамет несе летальні перебудови. Наприклад, у разі реципрокної транслокації зазвичай виживає не більше 50 % зигот.
Прикладом транслокації може служити «сімейний» синдром Дауна. При цьому захворюванні в одного з батьків фенотиопово не проявляються транслокація двадцять першої хромосоми на 14-ту. У такої людини з вірогідністю в 1/4 утворюються гамети з двома 21 хромосомами (одна вільна і одна транслокована). При злитті такої гамети з нормальною утворюється трисомік по 21 хромосомі.
Інший приклад — транслокація типу «Філадельфійська транслокація» між дев'ятою і двадцять другою хромосомами. У 95% випадків саме ця мутація є причиною хронічного мієлоцитарного лейкозу (англ. chronic myelogenous leukemia).
Робертсонівські транслокації, можливо, є причиною відмінностей між числом хромосом у близькоспоріднених видів. Показано, що два плеча 2-ї хромосоми людини відповідають 12 і 13 хромосомам шимпанзе. Можливо, 2-а хромосома утворилася в результаті робертсонівських транслокацій двох хромосом мавпоподібних предків людини. Таким же чином пояснюють той факт, що різні види дрозофіли мають від 3 до 6 хромосом.
Робертсонівські транслокації призвели до появи в Європі кількох видів-двійників (хромосомні раси) у мишей групи видів Mus musculus, які, як правило, географічно ізольовані один від одного. Набір і, як правило, експресія генів при робертсонівських транслокаціях не змінюються, тому види практично не відрізняються ззовні. Проте вони мають різні каріотипи, а плодючість при міжвидових схрещуваннях різко знижена.
Сайти ламкості
У 70-х роках XX століття було виявлено явище підвищеної ламкості хромосом — при фарбуванні метафазних хромосом культур клітин деяких індивідів барвниками деякі їхні ділянки залишалися безбарвними. Для цих ділянок характерна підвищена вірогідність хромосомних розривів. Природа цього явища ще не до кінця вивчена, можливо воно пов'язане з тим, що в цих ділянках хроматин знаходиться в неконденсованій формі. Дослідження говорять про зв'язок цього явища з однією з форм слабоумства (синдром Мартіна-Бела), а також захворюваністю рак.
Література
- Клаг У., Каммінгс М.Основи генетикию — М.: Світ, 2007.
- Біологія. Книга 1. Під ред. акад. РАМН Яригіна В. Н. — М.: Вища школа, 2003.
- Грін Н. та ін. Біологія — М.: Світ, 1990. — Т. 1—3.
- Жімулев І. Ф. Загальна та молекулярна генетика. — К.: Вид-во НГУ, 2003.
- Borgaonker, D.S. 1989. Chromosome variation in man: A catalogue of chromosomal variants and anomalies, 5th edition, New York: Alan R. Liss.
- Croce, C.M. 1996. The FHITgene at 3p14.2 is abnormal in lung cancer. Cell85:17-26.
- DeArce, M.A., and Kearns, A. 1984. The fragile X sindrome: The patients and their chromosomes. J. Med. Genet.21: 84-91.
- Gersh, M., et al. 1995. Evidence for a distinct region causing a catlike cry in patients with 5p delations. Am. J. Hum. Genet.56:1404-10.
- Hecht, F. 1988. Enigmatic fragile sites on human chromosomes. Trends Genet.4:121-22.
- Kaiser, P. 1984. Pericentric invertions: Problems and significance for clinical genetics. Hum. Genet.68:1-47.
- Lynch, M., and Conery, J.S. 2000. The evolutionary fate and consequences of duplicate genes. Science290: 1151-54.
- Lipski, J.R., Roth, J.R., and Weinstock, G.M. 1996. Chromosomal duplications in bactiria, fruit flies, and humans. Am. J. Hum. Genet.58:21-26.
- Madan, K. 1995. Paracentric invertions: a rewiew. Hum. Genet.96:503-515.
- Ohno, S. 1970. Evolution by gene duplication, New York: Springer-Verlag.
- Page, S.L., and Shaffer, L.G. 1997. Nonhomologous Robertsonian translocations form predominantly during female meiosis. Nature Genetics15:231-32.
Посилання
- Аберації хромосом // Велика українська енциклопедія : у 30 т. / проф. А. М. Киридон (відп. ред.) та ін. — 2016. — Т. 1 : А — Акц. — 592 с. — ISBN 978-617-7238-39-2.
