Черепашкові
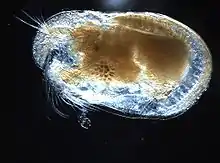
Черепа́шкові ра́ки[1] (іноді просто черепа́шкові[2]), або острако́ди[3] (лат. Ostracoda), — клас ракоподібних[4]. Дрібні ракоподібні з нечленистим тілом, зазвичай зжатим з боків, одягнений у двостулкову мушлею, із сімома парами придатків (антен, щелеп і ніг), із ногоподібними щупальцями верхніх щелеп, великими нижніми щелепами і нечленистим коротким черевцем. У викопному стані відомі з середнього/верхнього кембрія[5].
| Черепашкові | |
|---|---|
 | |
| Біологічна класифікація | |
| Царство: | Тварини (Animalia) |
| Тип: | Членистоногі (Arthropoda) |
| Підтип: | Ракоподібні (Crustacea) |
| Надклас: | Oligostraca |
| Клас: | Черепашкові (Ostracoda) Latreille, 1802 |
| Підкласи й ряди | |
Станом на 2008 рік, знайдено близько 2000 видів і 200 родів неморських черепашкових раків[6]. Однак, значна частина їх різноманіття все ще не описана, на що вказують незадокументовані гарячі точки біорізноманіття їх тимчасових місць проживання в Африці та Австралії[7]. Із відомого видового та родового різноманіття неморських остракод половина (1000 видів, 100 родів) належить до однієї родини (при загальній кількості — 13) — Cyprididae[7]. Багато Cyprididae зустрічаються в тимчасових водоймах і мають посухостійкі яйця, змішане/партеногенетичне розмноження та здатність плавати. Ці біологічні властивості попередньо адаптують їх до успішного поширення в цих середовищах існування[8].
Скам'янілості

Скам'янілості черепашкових раків на сьогоднішній день є найбільш поширеними скам'янілостями[9], віком починаючи з ордовицького періоду до наших часів. Основи схеми зони мікрофауни, яка спирається на форамініфер та черепашкових раків були сформовані Малкольмом Гартом[10]. Прісноводні черепашкові раки навіть були знайдені в балтійському бурштині еоценової епохи, імовірно, вони були занесені водою на дерева під час поводі[11].
Черепашкові раки особливо корисні для біостратиграфії морських товщ у місцевому або регіональному масшатбі, і є безцінними індикаторами палеоекології через їхню поширеність, невеликий розмір, невимогливість до умов зберігання, вилиняність, кальцифіковані двостулкові мушлі; стулки часто знаходять як мікроскам'янілості.
У Квінсленді 2013 року (оголошено у травні 2014) на місці відзначення двохсотріччя Австралії в місті Ріверслей на території Всесвітньої спадщини ЮНЕСКО, виявлено як чоловічі, так і жіночі рештки особин із дуже добре збереженими м'якими тканинами. Ця знахідка побила рекорд із Книги рекордів Гіннса як найстаріший пеніс[12]. У самців була виявлена найстаріша сперма, і при аналізі вона показала внутрішню структуру і була оцінена як найбільша сперма (на розмір тіла) серед усіх зареєстрованих тварин. Було досліджено, що скам'янілості досягли фосилізації за декілька днів завдяки фосфору в посліді кажанів у печері, де жили черепашкові раки[13].
Опис

Черепашкові раки — це невеликі ракоподібні, зазвичай розміром близько 1 мм, але варіюються від 0.2 до 30 мм, як у випадку з Gigantocypris. Їхні тіла сплющені побокам і захищені схожою на двостулкову хітинову або з умістом карбонату кальцію «мушлю». Замочна площадка (місце кріплення, англ. hinge) двох стулок знаходиться у верхній (дорсальній) частині тіла. Остракоди згруповані на основі загальної морфології. Хоча рання робота показала, що група не може бути монофілетичною[14]; і рання молекулярна філогенетика була неоднозначною щодо цього[15], нещодавні комбіновані аналізи молекулярних і морфологічних даних підтверджували монофілію в аналізах із ширшою вибіркою таксонів[16].
Тіло черепашкових раків оточене двостулковою хітиновою (іноді з умістом карбонату кальцію) мушлю, яка зовні нагадує мушлю молюска. Розрізняють стулку (тверді частини) і тіло з його придатками (м'які частини).
Частини тіла

Тіло складається з голови і грудей, розділених невеликим звуженням. На відміну від інших членистоногих, тіло чітко не поділене на сегменти. Черевце регресоване або відсутнє, тоді як гонади в дорослої особини відносно великі.
Голова є найбільшою частиною тіла і до неї прикріплені більшість придатків. Для плавання у воді використовуються 2 пари добре розвинених антен. Також у них присутні пара мандибул і 2 пари максил. На грудях зазвичай знаходяться 2 пари придатків, але вони зведені до однієї пари або зовсім відсутні у багатьох видів. Два виступи (rami) від кінчика хвоста спрямовані вниз і трохи вперед від задньої частини мушлі[17].
Черепашкові раки зазвичай не мають зябр, замість цього вони отримують кисень через зяброві пластини на поверхні тіла. У більшості також немає серця або кровоносної системи, і кров просто циркулює між стулками мушлі. Азотні відходи виводяться через залози на максилах і/або на антенах[17].
Основним видом відчуття черепашкових раків, імовірно, є дотик, оскільки вони мають кілька чутливих волосків на тілі та придатках. Однак у них є одне наупліарне око, а в деяких випадках і пара складних очей[17].
Екологія
Життєвий цикл
Самці черепашкових раків мають по два пеніси, відповідно до двох генітальних отворів (гонопорів) у самиць. Окремі сперматозоїди часто великі і згорнуті в сім'янниках перед спарюванням; в деяких випадках розгорнута сперма може бути в шість разів більшою за саму особину. Спарювання зазвичай відбувається під час того, як велика кількість самок скупчуються у рій і підпливають, щоб долучитися до самців. Деякі види повністю або частково партеногенетичні[17].
У більшості черепашкових раків яйця або відкладаються безпосередньо у воду як планктон, або прикріплюються до рослинності чи субстрату. Однак у деяких видів яйця покриті шкаралупою, що забезпечує їм більший ступінь захисту. Із яєць вилуплюються наупліуси, які вже мають тверду мушлю[17].
Хижаки
Різні хижаки полюють на черепашкових раків як у водному, так і наземному середовищі. Прикладом хижацтва в морському середовищі є деякі молюски родини Cuspidariidae, які виявляють черепашкових раків завдяки війкам, що виступають із вдихальних структур, звідки молюски втягують жертву за допомогою сильного всмоктування[18]. Хижацтво зі сторони вищих тварин теж існує; наприклад, амфібії, такі як жовтобрюхий тритон (Taricha granulosa), полюють на певних черепашкових[19].
Біолюмінісценція
Деякі черепашкові раки, такі як Vargula hilgendorfii, мають світловий орган, у якому вони виробляють люмінесцентні хімічні речовини[20]. Зазвичай світло слугує як захист від хижаків, хоча черепашкові раки в Карибському басейні використовують світло для спарювання. Таких черепашквоих раків називають «блакитний пісок» або «блакитні сльози», бо вони світяться блакитним у темряві. Їхні біолюмінісцентні властивості були важливими для Японії під час Другої світової війни, коли японська армія збирала їх у великій кількості з океану, щоб використовувати як зручне освітлення для читання карт та інших паперів уночі. Світло черепашкових раків, яке в Японії називали umihotaru, був достатнім для читання, але недостатньо яскравим, щоб бути помітним для ворога[21].
Примітки
- Черепашкові раки (Ostracoda). ВУЕ (укр.). Процитовано 3 січня 2022.
- https://website-designer-2149.business.site. КЛАС ЧЕРЕПАШКОВІ (OSTRACODA) - ПІДТИП ЗЯБРОДИШНІ (BRANCHIATA), АБО РАКОПОДІБНІ (CRUSTACEA) - ТИП ЧЛЕНИСТОНОГІ (ARTHROPODA) - ПІДРОЗДІЛ ЛИНЯЮЧІ (ECDYSOZOA) - РОЗДІЛ ТРИШАРОВІ (TRIPLOBLASTICA), або БІЛАТЕРАЛЬНІ (BILATERIA) ТВАРИНИ - ПІДЦАРСТВО СПРАВЖНІ БАГАТОКЛІТИННІ (EUMETAZOA) - ЦАРСТВО БАГАТОКЛІТИННІ (METAZOA) - ЗООЛОГІЯ БЕЗХРЕБЕТНИХ - Г. Й. Щербак - 2008. Библиотека биологических дисциплин (uk-ua). Процитовано 3 січня 2022.
- ОСТРАКОДИ | Геологічний словник. geodictionary.com.ua. Процитовано 3 січня 2022.
- WoRMS. Ostracoda. World Register of Marine Species. .
- Nicholas J. Butterfield, Maria I. Vélez, Thomas H. P. Harvey. Exceptionally preserved crustaceans from western Canada reveal a cryptic Cambrian radiation // Proceedings of the National Academy of Sciences. — National Academy of Sciences, 2012. — Vol. 109, iss. 5 (31 January). — ISSN 1091-6490 0027-8424, 1091-6490. — DOI:.
- K. Martens; I. Schon; C. Meisch; D. J. Horne (2008). Global diversity of ostracods (Ostracoda, Crustacea) in freshwater. Hydrobiologia 595 (1): 185–193. doi:10.1007/s10750-007-9245-4.
- K. Martens, S. A. Halse & I. Schon (2012). Nine new species of Bennelongia De Deckker & McKenzie, 1981 (Crustacea, Ostracoda) from Western Australia, with the description of a new subfamily. European Journal of Taxonomy 8: 1–56.
- Horne, D. J.; Martens, Koen (1998). An assessment of the importance of resting eggs for the evolutionary success of non-marine Ostracoda (Crustacea). У Brendonck, L.; De Meester, L.; Hairston, N. Evolutionary and ecological aspects of crustacean diapause 52. Advances in Limnology. с. 549–561. ISBN 9783510470549.
- David J. Siveter; Derek E. G. Briggs; Derek J. Siveter; Mark D. Sutton (2010). An exceptionally preserved myodocopid ostracod from the Silurian of Herefordshire, UK. Proceedings of the Royal Society B 277 (1687): 1539–1544. PMC 2871837. PMID 20106847. doi:10.1098/rspb.2009.2122.
- Malcolm B. Hart (1972). A correlation of the macrofaunal and microfaunal zonations of the Gault Clay in southeast England. У R. Casey; P. F. Rawson. Geological Journal (Special Issue 5): 267–288.
- Noriyuki Ikeya, Akira Tsukagoshi; David J. Horne (2005). Preface: The phylogeny, fossil record and ecological diversity of ostracod crustaceans. У Noriyuki Ikeya; Akira Tsukagoshi; David J. Horne. Hydrobiologia 538 (1–3): vii–xiii. doi:10.1007/s10750-004-4914-z.
- Oldest penis:
The oldest fossilised penis discovered to date dates back around 100 million years. It belongs to a crustacean called an ostracod, discovered in Brazil and measuring just 1mm across. - World's oldest sperm 'preserved in bat poo', Anna Salleh, Australian Broadcasting Corporation Science, 14 May 2014, accessed 15 May 2014
- Richard A. Fortey; Richard H. Thomas (1998). Arthropod Relationships. Chapman & Hall. ISBN 978-0-412-75420-3.
- S. Yamaguchi; K. Endo (2003). Molecular phylogeny of Ostracoda (Crustacea) inferred from 18S ribosomal DNA sequences: implication for its origin and diversification. Marine Biology 143 (1): 23–38. doi:10.1007/s00227-003-1062-3.
- Zaharoff, Alexander K.; Lindgren, Annie R.; Wolfe, Joanna M.; Oakley, Todd H. (1 січня 2013). Phylotranscriptomics to Bring the Understudied into the Fold: Monophyletic Ostracoda, Fossil Placement, and Pancrustacean Phylogeny. Molecular Biology and Evolution (англ.) 30 (1): 215–233. ISSN 0737-4038. PMID 22977117. doi:10.1093/molbev/mss216.
- Robert D. Barnes (1982). Invertebrate Zoology. Philadelphia: Holt-Saunders International. с. 680–683. ISBN 978-0-03-056747-6.
- John D. Gage; Paul A. Tyler (28 вересня 1992). Deep-Sea Biology: A Natural History of Organisms at the Deep-Sea Floor. University of Southampton. ISBN 978-0-521-33665-9. Проігноровано невідомий параметр
|name-list-style=(довідка) - C. Michael Hogan (2008). Rough-skinned Newt ("Taricha granulosa"). Globaltwitcher, ed. N. Stromberg. Архів оригіналу за 27 травня 2009.
- Osamu Shimomura (2006). The ostracod Cypridina (Vargula) and other luminous crustaceans. Bioluminescence: Chemical Principles and Methods. World Scientific. с. 47–89. ISBN 978-981-256-801-4.
- Jabr, Ferris. The Secret History of Bioluminescence. Hakai Magazine. Процитовано 6 липня 2016.