В-клітина пам'яті
В імунології В-клітина пам'яті (Memory B-cell, МВС "ем-би-си") — це тип В-лімфоцитів, що є частиною адаптивної імунної системи. Ці клітини розвиваються в зародкових центрах вторинних лімфоїдних органів. MBC циркулюють у кровотоці в стані спокою, іноді протягом десятиліть.[1] Їх функція полягає в тому, щоб "запам’ятати" характеристики антигену, який активував їх батьківську В-клітину під час початкової інфекції. Якщо MBC пізніше зустрінеться з тим самим антигеном, вона викликає прискорену, потужну та надійну вторинну імунну відповідь.[2] [3] На своїй мембрані MBC несуть В-клітинні рецептори (BCR, "бі-сі-ар"), ідентичні рецепторам їхньої батьківської клітини, які дозволяють їм розпізнавати антиген і створювати специфічну відповідь антитілами. [4]
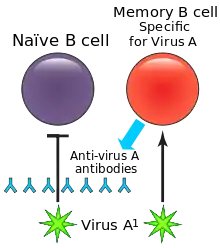
Розвиток та активація
Т-клітинно-залежні механізми
На шляху розвитку, залежного від Т-клітин, наївні (неактивні) фолікулярні В-клітини активуються антигенпрезентувальними фолікулярними Т-хелперами (TFH) під час початкової інфекції або первинної імунної відповіді.[3] Наївні В-клітини циркулюють через фолікули у вторинних лімфоїдних органах (тобто селезінці та лімфатичних вузлах ), де вони можуть бути активовані вільно циркулюючим чужорідним пептидом, що надходить через лімфу, або антигеном, представленим антигенпрезентувальними клітинами (АПК), такими як дендритні клітини (ДК).[5] В-клітини також можуть бути активовані шляхом зв’язування чужорідного антигену на периферії, звідки вони потім переміщуються у вторинні лімфоїдні органи.[3] Сигнал, від зв’язування пептиду з В-клітиною, змушує клітини мігрувати до краю фолікула, що межує з Т-клітинною зоною.[5]
В-клітини поглинають чужорідні пептиди, розщеплюють їх і експресують на головному комплексі гістосумісності класу II (MHCII), який є білком клітинної поверхні. У вторинних лімфоїдних органах більшість В-клітин потрапляє у В-клітинні фолікули, де утворюється зародковий центр. Більшість В-клітин зрештою диференціюється в плазматичні клітини або в MBC в зародковому центрі.[3] [6] TFHs, які експресують TCR, споріднені з пептидом (тобто специфічні для комплексу пептид-MHCII) на межі В-клітинного фолікула та Т-клітинної зони, зв’язуються з лігандом MHCII. Потім Т-клітини експресують молекулу ліганду CD40 (CD40L) і починають виділяти цитокіни, які спричиняють проліферацію В-клітин і перемикання класів імуноглобулінів, мутацію в генетичному коді В-клітин, яка змінює їх тип імуноглобуліну.[7] [8] Перемикання класів дозволяє майбутнім MBC секретувати різні типи антитіл у майбутніх імунних реакціях. [3] Потім В-клітини або диференціюються на плазматичні клітини, В-клітини зародкового центру або на MBC залежно від експресованих факторів транскрипції. Активовані В-клітини, які експресують транскрипційний фактор Bcl-6, потраплять у В-клітинні фолікули де відбуваються реакції зародкового центру.[7]
Потрапляючи в зародковий центр, В-клітини зазнають проліферації, за якою слідує мутація ділянки генетичного коду їх BCR, процес, відомий як соматична гіпермутація.[3] Мутації збільшують або зменшують спорідненість поверхневого рецептора до певного антигену - прогресивний процес, який називається дозріванням спорідненості. Після отримання цих мутацій рецептори на поверхні В-клітини "перевіряються" в зародковому центрі на їх спорідненість з поточним антигеном.[9] Клони В-клітини з мутаціями, які підвищили спорідненість їх поверхневих рецепторів, отримують сигнали про виживання через взаємодію зі спорідненими клітинами TFH.[2] [3] [10] В-клітини , які не мають достатньо високої спорідненості для отримання цих сигналів, а також В-клітини , які є потенційно аутореактивними, гинуть через апоптоз (негативний відбір).[6] Ці процеси збільшують мінливість у ділянках зв’язування антигену, таким чином кожна новостворена В-клітина має унікальний рецептор.[11]
Після диференціювання MBC переміщуються на периферію організму, де вони з більшою ймовірністю зустрінуть антиген у разі майбутнього контакту.[6] [2] [3] Багато циркулюючих MBC зосереджуються в ділянках тіла, які мають високу ймовірність контакту з антигеном, наприклад, у Пеєрових бляшках.
Процес диференціювання MBC в зародковому центрі ще не повністю вивчений.[3] Деякі дослідники припускають, що він відбувається випадково.[6] [4] Інші гіпотези припускають, що фактор транскрипції NF-κB і цитокін IL-24 беруть участь у процесі диференціювання MBC. [11] [3] Окрема гіпотеза стверджує, що В-клітини з відносно нижчою спорідненістю до антигену стають MBC, на відміну від В-клітин з відносно вищою спорідненістю, які стануть плазматичними клітинами.
Т-клітино-незалежні механізми
Не всі В-клітини, присутні в організмі, проходять соматичні гіпермутації. IgM+ MBC, які не зазнали перемикання класів імуноглобулінів, показують, що MBC можуть вироблятися незалежно від зародкових центрів.
Первинна реакція
Після інфікування багато В-клітин диференціюються в плазматичні клітини (також відомі як ефекторні В-клітини) які виробляють першу хвилю захисних антитіл і допомагають усунути інфекцію.[6] [2] Плазматичні клітини виділяють антитіла, специфічні для патогенів, але вони не можуть реагувати на повторне інфікування. Частина В-клітин з BCR, споріднених з антигеном, диференціюється в MBC, які довго зберігаються в організмі.[12] MBC можуть підтримувати експресію своїх BCR і зможуть швидко реагувати на повторну зустріч з патогеном.[6]
Вторинна реакція і пам'ять
MBC, що утворюються під час первинної імунної відповіді, є специфічними до антигену, який брав у цьому участь. При вторинній відповіді відповідатимуть MBC, специфічні до антигену або подібних антигенів.[3] Коли MBC знову зустрічають свій специфічний антиген, вони проліферують і диференціюються в плазматичні клітини, які потім реагують на антиген і видаляють його.[3] MBC, які в цей момент не диференціюються в плазматичні клітини, можуть знову увійти в зародкові центри для подальшого перемикання класів імуноглобулінів або соматичної гіпермутації для подальшого дозрівання спорідненості.[3] Диференціація MBC в плазматичні клітини відбувається набагато швидше, ніж диференціювання наївних В-клітин, що дозволяє їм виробляти більш ефективну вторинну імунну відповідь.[4] Ефективність і накопичення реакції MBC є основою дії вакцин і ревакцинацій.[4] [3]
Тривалість життя
В-клітини пам’яті можуть виживати десятиліттями, що дає їм здатність реагувати на багаторазовий вплив одного і того ж антигену.[3] Вважається, що довготривале виживання є результатом дії певних антиапоптозних генів, які більш високо експресуються у MBC, ніж у інших підгрупах В-клітин.[6] Крім того для довготривалого виживання, MBC не потребує постійної взаємодії ні з антигеном, ні з Т-клітинами.[4]
Маркери
MBC зазвичай розрізняють за маркером клітинної поверхні CD27, хоча деякі підгрупи його не експресують. MBC, які не мають CD27, як правило, пов’язані з виснаженими В-клітинами або певними аутоімунними станами, такими як ВІЛ, вовчак або ревматоїдний артрит.[2] [3]
Оскільки у В-клітинах зазвичай відбувається перемикання класів імуноглобулінів, вони можуть експресувати цілу низку антитіл. Деякі специфічні властивості окремих антитіл описані нижче:
- IgM: MBC, які експресують IgM, зосереджуються в мигдаликах, Пеєрових бляшках та лімфатичних вузлах.[3] Ця підгрупа MBC з більшою ймовірністю зазнає проліферації та знову потрапить у зародковий центр під час вторинної імунної відповіді. [4]
- IgG: MBC, які експресують IgG, зазвичай диференціюються в плазматичні клітини.[4]
- IgE: MBC, які експресують IgE, дуже рідкісні у здорових людей. Це може статися тому, що В-клітини, які експресують IgE, частіше диференціюються в плазматичні клітини, а не в В-клітини пам'яті [4]
- IgD: MBC, які експресують тільки IgD, зустрічаються дуже рідко, вони зосереджені в мигдаликах. [13]
Рецептор CCR6, як правило, є маркером В-клітин, які в кінцевому підсумку диференціюються в MBC. Цей рецептор реагує на хемокіни, які є хімічними посередниками направленого руху В-клітин. MBC можуть мати цей рецептор, що дозволяє їм переміщатися із зародкового центру в тканини, де вони мають більшу ймовірність зіткнутися з антигеном.[6]
Підгрупи
MBC незалежні від зародкового центру
Ця підгрупа клітин диференціюється з активованих В-клітин у MBC перед тим, як потрапити в зародковий центр. В-клітини, які мають високий рівень взаємодії з TFH в межах В-клітинного фолікула, мають більшу схильність до проникнення в зародковий центр. В-клітини, які розвиваються у MBC незалежно від зародкових центрів, ймовірно, зазнають передачу сигналів CD40 і цитокінів від Т-клітин.[14] Перемикання класів імуноглобулінів все ще може відбуватися до взаємодії із зародковим центром, тоді як соматична гіпермутація відбувається лише після взаємодії із зародковим центром.[14] Відсутність соматичної гіпермутації вважається корисною; нижчий рівень дозрівання спорідненості означає, що ці В-клітини пам’яті менш спеціалізовані до певного антигену і можуть розпізнавати більш широкий спектр антигенів. [11] [15] [4]
MBC незалежні від Т-клітин
В-клітини незалежні від Т-клітин є субгрупою, яка називається клітинами В1. Ці клітини зазвичай знаходяться в очеревній порожнині. При повторному введенні антигену деякі з цих клітин В1 можуть диференціюватися в MBC без взаємодії з Т-клітинами. [4] Вони виробляють антитіла IgM. [16]
MBC T-bet
В-клітини T-bet є підгрупою, яка експресує фактор транскрипції T-bet. T-bet пов'язаний з перемиканням класів імуноглобулінів. Вважається, що В-клітини T-bet також мають важливе значення в імунній відповіді проти внутрішньоклітинних бактеріальних та вірусних інфекцій.[17]
Вакцинація
Дія вакцин заснована на уявленні про імунологічну пам'ять. Профілактична ін'єкція непатогенного антигену в організм дозволяє йому сформувати стійку імунологічну пам'ять . Ін’єкція антигену призводить до реакції появи антитіл з подальшим виробленням В-клітин пам’яті. Ці В-клітини пам’яті швидко реактивуються після інфікування відповідним патогеном і можуть ефективно захищати організм від хвороби.[18]
Дивись також
Посилання
- Cutting edge: long-term B cell memory in humans after smallpox vaccination. Journal of Immunology 171 (10): 4969–73. November 2003. PMID 14607890. doi:10.4049/jimmunol.171.10.4969. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Memory B Cells of Mice and Humans. Annual Review of Immunology 35 (1): 255–284. April 2017. PMID 28142324. doi:10.1146/annurev-immunol-041015-055531. Проігноровано невідомий параметр
|vauthors=(довідка) - Human memory B cells. Leukemia 30 (12): 2283–2292. December 2016. PMID 27499139. doi:10.1038/leu.2016.226. Проігноровано невідомий параметр
|vauthors=(довідка) - Memory B cells. Nature Reviews. Immunology 15 (3): 149–59. March 2015. PMID 25677494. doi:10.1038/nri3802. Проігноровано невідомий параметр
|vauthors=(довідка) - Visualization of specific B and T lymphocyte interactions in the lymph node. Science 281 (5373): 96–9. July 1998. PMID 9651253. doi:10.1126/science.281.5373.96. Проігноровано невідомий параметр
|vauthors=(довідка) - Plasma cell and memory B cell differentiation from the germinal center. Current Opinion in Immunology. Lymphocyte development and activation * Tumour immunology 45: 97–102. April 2017. PMID 28319733. doi:10.1016/j.coi.2017.03.006. Проігноровано невідомий параметр
|vauthors=(довідка) - Heterogeneity in the differentiation and function of memory B cells. Trends in Immunology 33 (12): 590–7. December 2012. PMC 3505266. PMID 22920843. doi:10.1016/j.it.2012.07.005. Проігноровано невідомий параметр
|vauthors=(довідка) - Phan, Tri Giang; Tangye, Stuart G (1 квітня 2017). Memory B cells: total recall. Current Opinion in Immunology. Lymphocyte development and activation * Tumour immunology (англ.) 45: 132–140. ISSN 0952-7915. doi:10.1016/j.coi.2017.03.005.
- The continuing story of T-cell independent antibodies. Immunological Reviews 288 (1): 128–135. March 2019. PMC 6653682. PMID 30874357. doi:10.1111/imr.12754. Проігноровано невідомий параметр
|vauthors=(довідка) - Germinal centers. Annual Review of Immunology 30 (1): 429–57. 26 березня 2012. PMID 22224772. doi:10.1146/annurev-immunol-020711-075032. Проігноровано невідомий параметр
|vauthors=(довідка) - Regulation of memory B and plasma cell differentiation. Current Opinion in Immunology. Lymphocyte development and activation * Tumour immunology 45: 126–131. April 2017. PMID 28359033. doi:10.1016/j.coi.2017.03.003. Проігноровано невідомий параметр
|vauthors=(довідка) - The germinal center reaction. The Journal of Allergy and Clinical Immunology 126 (5): 898–907; quiz 908–9. November 2010. PMID 21050940. doi:10.1016/j.jaci.2010.09.007. Проігноровано невідомий параметр
|vauthors=(довідка) - Human memory B cells. Leukemia 30 (12): 2283–2292. December 2016. PMID 27499139. doi:10.1038/leu.2016.226. Проігноровано невідомий параметр
|vauthors=(довідка) - Memory B cells. Nature Reviews. Immunology 15 (3): 149–59. March 2015. PMID 25677494. doi:10.1038/nri3802. Проігноровано невідомий параметр
|vauthors=(довідка) - An antigen to remember: regulation of B cell memory in health and disease. Current Opinion in Immunology. Lymphocyte development and activation * Tumour immunology 45: 89–96. April 2017. PMC 7126224. PMID 28319732. doi:10.1016/j.coi.2017.03.004. Проігноровано невідомий параметр
|vauthors=(довідка) - B-1 B cell development in the fetus and adult. Immunity 36 (1): 13–21. January 2012. PMC 3269035. PMID 22284417. doi:10.1016/j.immuni.2011.11.017. Проігноровано невідомий параметр
|vauthors=(довідка) - T-bet+ memory B cells: Generation, function, and fate. Immunological Reviews 288 (1): 149–160. March 2019. PMC 6626622. PMID 30874358. doi:10.1111/imr.12736. Проігноровано невідомий параметр
|vauthors=(довідка) - The geography of memory B cell reactivation in vaccine-induced immunity and in autoimmune disease relapses. Immunological Reviews 296 (1): 62–86. July 2020. PMID 32472583. doi:10.1111/imr.12862. Проігноровано невідомий параметр
|vauthors=(довідка)