Рецептор інсуліноподібного фактору росту 2 типу
Рецептор інсуліноподібного фактору росту 2 типу (англ. Insulin like growth factor 2 receptor, IGF2R), також катіон-незалежний рецептор маноза-6-фосфату (англ. cation-independent mannose-6-phosphate receptor, CI-MPR) – білок, який кодується геном IGF2R, розташованим у людей на довгому плечі 6-ї хромосоми.[4][5][6]. Довжина поліпептидного ланцюга білка становить 2 491 амінокислот, а молекулярна маса — 274 375[7]. IGF2R є багатофункціональним рецептором, який зв'язується з інсуліноподібним фактором росту 2 типу на поверхні клітини та маноза-6-фосфатом М6Р-мічених білків в транс-Гольджі[6].
| 10 | 20 | 30 | 40 | 50 | ||||
|---|---|---|---|---|---|---|---|---|
| MGAAAGRSPH | LGPAPARRPQ | RSLLLLQLLL | LVAAPGSTQA | QAAPFPELCS | ||||
| YTWEAVDTKN | NVLYKINICG | SVDIVQCGPS | SAVCMHDLKT | RTYHSVGDSV | ||||
| LRSATRSLLE | FNTTVSCDQQ | GTNHRVQSSI | AFLCGKTLGT | PEFVTATECV | ||||
| HYFEWRTTAA | CKKDIFKANK | EVPCYVFDEE | LRKHDLNPLI | KLSGAYLVDD | ||||
| SDPDTSLFIN | VCRDIDTLRD | PGSQLRACPP | GTAACLVRGH | QAFDVGQPRD | ||||
| GLKLVRKDRL | VLSYVREEAG | KLDFCDGHSP | AVTITFVCPS | ERREGTIPKL | ||||
| TAKSNCRYEI | EWITEYACHR | DYLESKTCSL | SGEQQDVSID | LTPLAQSGGS | ||||
| SYISDGKEYL | FYLNVCGETE | IQFCNKKQAA | VCQVKKSDTS | QVKAAGRYHN | ||||
| QTLRYSDGDL | TLIYFGGDEC | SSGFQRMSVI | NFECNKTAGN | DGKGTPVFTG | ||||
| EVDCTYFFTW | DTEYACVKEK | EDLLCGATDG | KKRYDLSALV | RHAEPEQNWE | ||||
| AVDGSQTETE | KKHFFINICH | RVLQEGKARG | CPEDAAVCAV | DKNGSKNLGK | ||||
| FISSPMKEKG | NIQLSYSDGD | DCGHGKKIKT | NITLVCKPGD | LESAPVLRTS | ||||
| GEGGCFYEFE | WHTAAACVLS | KTEGENCTVF | DSQAGFSFDL | SPLTKKNGAY | ||||
| KVETKKYDFY | INVCGPVSVS | PCQPDSGACQ | VAKSDEKTWN | LGLSNAKLSY | ||||
| YDGMIQLNYR | GGTPYNNERH | TPRATLITFL | CDRDAGVGFP | EYQEEDNSTY | ||||
| NFRWYTSYAC | PEEPLECVVT | DPSTLEQYDL | SSLAKSEGGL | GGNWYAMDNS | ||||
| GEHVTWRKYY | INVCRPLNPV | PGCNRYASAC | QMKYEKDQGS | FTEVVSISNL | ||||
| GMAKTGPVVE | DSGSLLLEYV | NGSACTTSDG | RQTTYTTRIH | LVCSRGRLNS | ||||
| HPIFSLNWEC | VVSFLWNTEA | ACPIQTTTDT | DQACSIRDPN | SGFVFNLNPL | ||||
| NSSQGYNVSG | IGKIFMFNVC | GTMPVCGTIL | GKPASGCEAE | TQTEELKNWK | ||||
| PARPVGIEKS | LQLSTEGFIT | LTYKGPLSAK | GTADAFIVRF | VCNDDVYSGP | ||||
| LKFLHQDIDS | GQGIRNTYFE | FETALACVPS | PVDCQVTDLA | GNEYDLTGLS | ||||
| TVRKPWTAVD | TSVDGRKRTF | YLSVCNPLPY | IPGCQGSAVG | SCLVSEGNSW | ||||
| NLGVVQMSPQ | AAANGSLSIM | YVNGDKCGNQ | RFSTRITFEC | AQISGSPAFQ | ||||
| LQDGCEYVFI | WRTVEACPVV | RVEGDNCEVK | DPRHGNLYDL | KPLGLNDTIV | ||||
| SAGEYTYYFR | VCGKLSSDVC | PTSDKSKVVS | SCQEKREPQG | FHKVAGLLTQ | ||||
| KLTYENGLLK | MNFTGGDTCH | KVYQRSTAIF | FYCDRGTQRP | VFLKETSDCS | ||||
| YLFEWRTQYA | CPPFDLTECS | FKDGAGNSFD | LSSLSRYSDN | WEAITGTGDP | ||||
| EHYLINVCKS | LAPQAGTEPC | PPEAAACLLG | GSKPVNLGRV | RDGPQWRDGI | ||||
| IVLKYVDGDL | CPDGIRKKST | TIRFTCSESQ | VNSRPMFISA | VEDCEYTFAW | ||||
| PTATACPMKS | NEHDDCQVTN | PSTGHLFDLS | SLSGRAGFTA | AYSEKGLVYM | ||||
| SICGENENCP | PGVGACFGQT | RISVGKANKR | LRYVDQVLQL | VYKDGSPCPS | ||||
| KSGLSYKSVI | SFVCRPEARP | TNRPMLISLD | KQTCTLFFSW | HTPLACEQAT | ||||
| ECSVRNGSSI | VDLSPLIHRT | GGYEAYDESE | DDASDTNPDF | YINICQPLNP | ||||
| MHGVPCPAGA | AVCKVPIDGP | PIDIGRVAGP | PILNPIANEI | YLNFESSTPC | ||||
| LADKHFNYTS | LIAFHCKRGV | SMGTPKLLRT | SECDFVFEWE | TPVVCPDEVR | ||||
| MDGCTLTDEQ | LLYSFNLSSL | STSTFKVTRD | SRTYSVGVCT | FAVGPEQGGC | ||||
| KDGGVCLLSG | TKGASFGRLQ | SMKLDYRHQD | EAVVLSYVNG | DRCPPETDDG | ||||
| VPCVFPFIFN | GKSYEECIIE | SRAKLWCSTT | ADYDRDHEWG | FCRHSNSYRT | ||||
| SSIIFKCDED | EDIGRPQVFS | EVRGCDVTFE | WKTKVVCPPK | KLECKFVQKH | ||||
| KTYDLRLLSS | LTGSWSLVHN | GVSYYINLCQ | KIYKGPLGCS | ERASICRRTT | ||||
| TGDVQVLGLV | HTQKLGVIGD | KVVVTYSKGY | PCGGNKTASS | VIELTCTKTV | ||||
| GRPAFKRFDI | DSCTYYFSWD | SRAACAVKPQ | EVQMVNGTIT | NPINGKSFSL | ||||
| GDIYFKLFRA | SGDMRTNGDN | YLYEIQLSSI | TSSRNPACSG | ANICQVKPND | ||||
| QHFSRKVGTS | DKTKYYLQDG | DLDVVFASSS | KCGKDKTKSV | SSTIFFHCDP | ||||
| LVEDGIPEFS | HETADCQYLF | SWYTSAVCPL | GVGFDSENPG | DDGQMHKGLS | ||||
| ERSQAVGAVL | SLLLVALTCC | LLALLLYKKE | RRETVISKLT | TCCRRSSNVS | ||||
| YKYSKVNKEE | ETDENETEWL | MEEIQLPPPR | QGKEGQENGH | ITTKSVKALS | ||||
| SLHGDDQDSE | DEVLTIPEVK | VHSGRGAGAE | SSHPVRNAQS | NALQEREDDR | ||||
| VGLVRGEKAR | KGKSSSAQQK | TVSSTKLVSF | HDDSDEDLLH | I |
Структура гену
Ген IGF2R містить 48 екзонів і охоплює область розміром близько 136 тисяч пар основ[8]. Встановлено, що промоторні області генів IGF2R миші та людини розташовані в межах CpG-острівців, де відсутні ТАТА- і СААТ-бокси. Деякі регуляторні елементи зберігаються в миші і людини, в тому числі 3 E-бокси та і кілька GC-боксів. Другий інтрон гену IGF2R як у миші, так і у людини містить другий CpG-острівець[9].
На сьогодні для IGF2R відомо 9 однонуклеотидних поліморфізмів та 3 варіанти амінокислот в ліганд-зв'язуючому домені M6P/IGF2R, які можуть під впливом відбору в організмі людини[10] .
Модель геномного імпринтингу IGF2R
Ген IGF2R є імпринтованим, тобто його експресія здійснюється лише з локусу хромосоми одного із батьків. Експресія цього гену (в людини) відбувається на довгому плечі 6-ї хромосоми материнського походження у позиції 26-27 й регулюється за допомогою нкРНК (англ. ncRNA). При цьому експресія гену, що відповідає за синтез некодуючої РНК, відбувається з локусу хромосоми батьківського походження. Геномна організація і патерн імпринтингу кластера Igf2r миші лише частково зберігається в синтенній області людини[11].
У регуляції імпринтованого локусу цього гену у миші задіяні диференційне метилювання CpG-динуклеотидів у другому інтроні й антизмістовна некодуюча РНК Air[12]. Таким чином, на хромосомі материнського походження, ген Air репресований (зумовлено гіперметилюванням ICE (англ. imprint control element), що розташований в його промоторі[11]), а три білок-кодуючі гени, що знаходяться під його негативним контролем — активні. На хромосомі батьківського походження послідовність ДНК в області ICE є неметильована, що обумовлює транскрипцію з батьківської хромосоми нкРНК Air розміром 108 т. н., яка забезпечує сайленсинг алелей батьківського походження як самого гена IGF2R, з яким він частково перекривається, так і генів SLC22A2 і SLC22A3, з якими він не перекривається[13].
У сумчастих Igf2r також імпринтований, проте експресія материнського алеля здійснюється без участі диференційного метилювання, через відсутность CpG-острівця в другому інтроні й і антизмістовної транскрипції[12].
Структура IGF2R
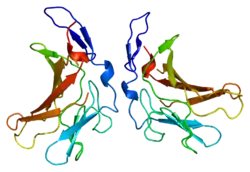
За структурою IGF2R відноситься до трансмембранних білків I типу (тобто, він має один трансмембранний домен з його С-кінцем на цитоплазматичній стороні ліпіднийого бішару) з великим позаклітинним/просвітним (просвіт транс-сітки апарату Гольджі) доменом та відносно коротким цитоплазматичним хвостом[14]. Позаклітинний домен містить невелику область, що гомологічна колаген-зв'язуючому доменому фибронектину і п'ятнадцять повторів з приблизно 147 амінокислотних залишків. Кожен з цих повторів гомологічний 157 залишку позаклітинного домену рецептора манози 6-фосфату. Зв'язування з IGF2 опосередковано через один з повторів, в той час як два різних повтори відповідальні за зв'язування з маноза-6-фосфатом. Молекулярна маса IGF2R становить приблизно 300 кДа; він існує та функціонує як димер.
Фізіологічна роль
IGF2R здійснює очищення IGF2 від поверхні клітини для ослабленої сигналізації, а також транспортування попередників лізосомальних кислих гідролаз з апарату Гольджі в лізосоми. Після зв'язування IGF2 на поверхні клітини, молекули IGF2R накопичуються в клатринових везикулах, що формуються та поглинаються. У просвіті транс-сітки апарату Гольджі, IGF2R зв'язує М6Р-мічений «вантаж»[14]. Молекули IGF2R (зв'язані з їх «вантажем») впізнаються GGA родиною клатринових адаптерних білків та накопичуються в клатринових везикулах, що формуються[15]. Молекули IGF2R з поверхні клітин та апарату Гольджі поміщаються в ранні ендосоми де, при відносно низькому рН середовища ендосом, вони вивільняють свій «вантаж». Молекули IGF2R повертаються назад до апарату Гольджі за допомогою ретромерного комплексу, знову шляхом взаємодії з GGA та везикулами. Білковий «вантаж» поміщається в лізосоми за допомогою пізніх ендосом незалежно від IGF2R.
Інтерактом IGF2R
Було показано, що рецептор інсуліноподібного фактору росту 2 типу взаємодіє з маноза-6-фосфат-рецептор-зв'язуючим білком 1 (M6PRBP1, PLIN3)[16][17].
Особливості еволюції IGF2R
Рецептор інсуліноподібного фактору росту 2 типу еволюціонував від катіон-незалежного рецептора маноза-6-фосфату та уперше з'явився в однопрохідних. Сайт зв'язування IGF2 був, ймовірно, набутий випадково при утворенні екзонного сайта сплайсингу енхансерного кластера в екзоні 34 та ймовірно обумовлено вставками повторюваних елементів, довжиною в кілька тисяч пар основ, в попередньому інтроні. Шестиразове збільшення афінності в еволюції звірів збігається з виникненням імпринтингу і узгоджується з теорією батьківського конфлікту[18].
IGF2R та обдарованість
Відмінності в рівні інтелекту індивідуумів в значній мірі визначаються відмінностями в генах. У 90-х роках американський вчений XX століття Річард Пломін показав, що більшість дітей з дуже високим IQ мають «ген інтелекту» IGF2R який кодує рецептор для так званого інсуліноподібного фактору росту 2 типу. Проте у 2009 році британські вчені під керівництвом Яна Дірі показали, що людський інтелект досить полігенний (визначається багатьма генами)[19].
Див. також
Примітки
- Захворювання, генетично пов'язані з Рецептор інсуліноподібного фактору росту 2 типу переглянути/редагувати посилання на ВікіДаних.
- Human PubMed Reference:.
- Mouse PubMed Reference:.
- HUGO Gene Nomenclature Commitee, HGNC:5467 (англ.). Процитовано 12 лютого 2018.
- Oshima A, Nolan CM, Kyle JW, Grubb JH, Sly WS (February 1988). The human cation-independent mannose 6-phosphate receptor. Cloning and sequence of the full-length cDNA and expression of functional receptor in COS cells. J. Biol. Chem. 263 (5): 2553–62. PMID 2963003.
- Laureys G, Barton DE, Ullrich A, Francke U (October 1988). Chromosomal mapping of the gene for the type II insulin-like growth factor receptor/cation-independent mannose 6-phosphate receptor in man and mouse. Genomics 3 (3): 224–9. PMID 2852162. doi:10.1016/0888-7543(88)90083-3.
- UniProt, P11717 (англ.). Процитовано 12 лютого 2018.
- Killian, J. K., Jirtle, R. L. Genomic structure of the human M6P/IGF2 receptor. Mammalian Genome 10: 74-77, 1999.
- Smrzka, O. W., Fae, I., Stoger, R., Kurzbauer, R., Fischer, G. F., Henn, T., Weith, A., Barlow, D. P. Conservation of a maternal-specific methylation signal at the human IGF2R locus. Hum. Molec. Genet. 4: 1945-1952, 1995.
- Killian, J. K., Oka, Y., Jang, H.-S., Fu, X., Waterland, R. A., Sohda, T., Sakaguchi, S., Jirtle, R. L. Mannose 6-phosphate/insulin-like growth factor 2 receptor (M6P/IGF2R) variants in American and Japanese populations. Hum. Mutat. 18: 25-31, 2001.
- Эпигенетика эмбрионального развития человека. 6.2 Модели импринтинга. 16.03.2015
- Медицинский портал Казахстана ODK. Регуляция импринтированного статуса. 06.04.2014. Архів оригіналу за 01.11.2016. Процитовано 31.10.2016.
- Медицинский портал Казахстана ODK. Обеспечение сайленсинга генов в локусе. 16.03.2014. Архів оригіналу за 01.11.2016. Процитовано 31.10.2016.
- Ghosh P, Dahms NM, Kornfeld S (March 2003). Mannose 6-phosphate receptors: new twists in the tale. Nat. Rev. Mol. Cell Biol. 4 (3): 202–12. PMID 12612639. doi:10.1038/nrm1050.
- Ghosh P, Kornfeld S (July 2004). The GGA proteins: key players in protein sorting at the trans-Golgi network. Eur. J. Cell Biol. 83 (6): 257–62. PMID 15511083. doi:10.1078/0171-9335-00374.
- Díaz E, Pfeffer SR (May 1998). TIP47: a cargo selection device for mannose 6-phosphate receptor trafficking. Cell 93 (3): 433–43. PMID 9590177. doi:10.1016/S0092-8674(00)81171-X.
- Orsel JG, Sincock PM, Krise JP, Pfeffer SR (August 2000). Recognition of the 300-kDa mannose 6-phosphate receptor cytoplasmic domain by 47-kDa tail-interacting protein. Proc. Natl. Acad. Sci. U.S.A. 97 (16): 9047–51. PMC 16819. PMID 10908666. doi:10.1073/pnas.160251397.
- Williams C, Hoppe HJ, Rezgui D, Strickland M, Forbes BE, Grutzner F, Frago S, Ellis RZ, Wattana-Amorn P, Prince SN, Zaccheo OJ, Nolan CM, Mungall AJ, Jones EY, Crump MP, Hassan AB (November 2012). Exon splice enhancer primes IGF2:IGF2R binding site structure and function evolution. Science 338 (6111): 1209–1213. PMID 23197533. doi:10.1126/science.1228633.
- Wlodzimierz Klonowski (2015). Genius and nonlinear dynamics. EPJ Nonlinear Biomedical Physics 3:7
Література
- Killian J.K., Jirtle R.L. (1999). Genomic structure of the human M6P/IGF2 receptor.. Mamm. Genome 10: 74 — 77. PubMed DOI:10.1007/s003359900947
- Puertollano R., Aguilar R.C., Gorshkova I., Crouch R.J., Bonifacino J.S. (2001). Sorting of mannose 6-phosphate receptors mediated by the GGAs.. Science 292: 1712 — 1716. PubMed DOI:10.1126/science.1060750
- Zhang H., Li X.-J., Martin D.B., Aebersold R. (2003). Identification and quantification of N-linked glycoproteins using hydrazide chemistry, stable isotope labeling and mass spectrometry.. Nat. Biotechnol. 21: 660 — 666. PubMed DOI:10.1038/nbt827
- Giorgianni F., Zhao Y., Desiderio D.M., Beranova-Giorgianni S. (2007). Toward a global characterization of the phosphoproteome in prostate cancer cells: identification of phosphoproteins in the LNCaP cell line.. Electrophoresis 28: 2027 — 2034. PubMed DOI:10.1002/elps.200600782
- Carrascal M., Ovelleiro D., Casas V., Gay M., Abian J. (2008). Phosphorylation analysis of primary human T lymphocytes using sequential IMAC and titanium oxide enrichment.. J. Proteome Res. 7: 5167 — 5176. PubMed DOI:10.1021/pr800500r
- Oshima A., Nolan C.M., Kyle J.W., Grubb J.H., Sly W.S. (1988). The human cation-independent mannose 6-phosphate receptor. Cloning and sequence of the full-length cDNA and expression of functional receptor in COS cells.. J. Biol. Chem. 263: 2553 — 2562. PubMed