Рибосомальний зсув рамки зчитування
Рибосомальний зсув рамки зчитування (англ. ribosomal frameshifting), також відомий як трансляційний зсув рамки зчитування (англ. translational frameshifting) або трансляційне перекодування (англ. translational recoding) — це біологічне явище, що виникає під час трансляції, й внаслідок якого продукується декілька унікальних білків з однієї мРНК[1]. Процес може бути запрограмований нуклеотидною послідовністю мРНК і іноді впливає на вторинну, тривимірну структуру мРНК[2]. Це явище описане здебільшого у вірусів (особливо у ретровірусів), ретротранспозонах, бактеріальних вставних елементах, а також у деяких клітинних генах[3].
Огляд процесу
Білки транслюються шляхом зчитування кодонів (трьох послідовно розташованих нуклеотидних залишків) з ланцюга мРНК, з одного кінця мРНК до іншого (від 5'- до 3'-кінця). Кожен кодон транслюється в одну амінокислоту. Таким чином, зсув будь-якої кількості нуклеотидів, що не ділиться на 3, в рамці зчитування, призведе до того, що наступні кодони будуть читатися по-іншому[4]. Це фактично змінює рибосомальну рамку зчитування. Наприклад, наступне речення, коли читається з початку, має сенс для читача:
|Старт|ТУТ АБО ТАМ БУВ ЛІС АБО САД ... |Старт|123 123 123 123 123 123 123 ...
Проте, зсув рамки зчитування між літерами T і У першого слова, тобто читання речення з другої літери першого слова (фактично «зсув рамки +1», якщо розглядати позицію 0 як початкову позицію літери Т) призведе до такого:
T|Старт|УТА БОТ АМБ УВЛ ІСА БОС АД... -|Старт|123 123 123 123 123 123 12...
Тепер речення не має сенсу. У разі транслювання рибосомою, зсув рамки зчитування може призвести або до нонсенсу (появи передчасного стоп-кодону) після зсуву рамки зчитування, або до створення повністю нового білка після зсуву рамки зчитування. У тому разі, коли зсув рамки зчитування призводить до нонсенс-мутації, НОР-шляхом (нонсенс-опосередкований розпад мРНК) може бути зруйнований мРНК-транскрипт, тому зсув рамки зчитування слугував би методом регуляції рівня експресії асоційованого гена[5] .
Функція
У вірусів це явище може бути запрограмоване відбуватися в певних ділянках і дозволяє вірусу кодувати кілька типів білків в одній і тій же мРНК. Яскравими прикладами слугують ВІЛ-1 (вірус імунодефіциту людини)[6], ВСР (вірус саркоми Рауса)[7] і вірус грипу (грип)[8], які всі покладаються на зсув рамки зчитування для створення правильного співвідношення білків 0-рамки (нормальна трансляція) та «транс-рамки» (кодуються послідовністю зі зсувом рамки зчитування). Його призначення у вірусів — це насамперед для вміщення більшої кількості генетичної інформації в меншій кількості генетичного матеріалу.
У еукаріотів, схоже, він відіграє певну роль у регуляції рівнів експресії генів шляхом генерування передчасних стоп-кодонів та створення нефункціональних транскриптів[3][9].
Типи зсуву рамок зчитування
Найпоширенішим типом зсув рамок зчитування є -1 зсув рамки зчитування або запрограмований -1 рибосомальний зсув рамки зчитування. Іншими, рідкіснішими типами зсувів рамок зчитування є +1 і -2 зсуви рамок зчитування[2]. −1 та +1, як вважається, контролюються різними механізмами, про які ідеться нижче. Обидва механізми керуються кінетично.
Запрограмований -1 рибосомальний зсув рамки зчитування
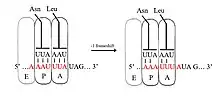
За -1 зсуву рамки зчитування, рибосома зісковзує на один нуклеотид назад і продовжує трансляцію в -1 рамці зчитування. Є, як правило, три елементи, що містять сигнал -1 зсуву рамки зчитування: слизька послідовність, спейсерна область та вторинна структура РНК. Слизька послідовність відповідає мотиву X_XXY_YYZ, де XXX є будь-якими трьома однаковими нуклеотидами (хоча трапляються деякі винятки), YYY це, як правило, УУУ або ААА, а Z — це А, Ц або У. Оскільки структура цього мотиву містить 2 прилеглих 3-нуклеотидні повтори, вважається, що -1 зсув рамки зчитування описується моделлю тандемного ковзання, за якого одночасно антикодон тРНК в рибосомальному Р-сайті переспаровується з XXY на XXX, а антикодон в А-сайті переспаровується з YYZ на YYY. Ці нові пари ідентичні парам 0-рамки зчитування, за винятком їхніх третіх позицій. Ця відмінність не має значного негативного впливу на зв'язування з антикодоном, тому що третій нуклеотид в кодоні, який перебуває в зміщеному (воблівському) положені, має слабшу специфічність зв'язування тРНК антикодону, ніж перший і другий нуклеотиди[2][10]. У цій моделі структура мотивів пояснюється тим, що антикодони в першому і другому положені повинні мати можливість ідеально спаровуватися як в 0, так і в -1 рамці зчитування. Таким чином, нуклеотиди 2 та 1 повинні бути ідентичними й нуклеотиди 3 та 2 також повинні бути ідентичними, що зумовлює необхідність у послідовності 3 однакових нуклеотидів для кожної тРНК, яка «ковзає»[11].
+1 рибосомальний зсув рамки зчитування
«Слизька послідовність» сигналу +1 зсуву рамки зчитування не має однакового мотиву, і натомість, здається, здійснюється паузою рибосоми на послідовності, що кодує рідкісну амінокислоту[12]. Рибосоми не транслюють білки з постійною швидкістю, незалежно від послідовності. Деякі кодони транслюються довше, тому що немає достатньої кількості тРНК саме цього кодону в цитозолі[13]. Через це затримання, існують невеликі ділянки послідовностей кодонів, які контролюють швидкість рибосомального зсуву рамки зчитування. Зокрема, рибосома повинна зробити паузу, щоб дочекатися прибуття рідкісної тРНК, і це збільшує кінетичну сприятливість рибосоми та асоційованої тРНК до проковзування на нову рамку зчитування[12][14]. За цієї моделі, зміна рамки зчитування спричинена проковзуванням однієї тРНК, а не двох.
Контролювальні механізми
Рибосомальний зсув рамки зчитування може контролюватися механізмами, знайденими в послідовності мРНК (цис-діючій). Зазвичай це стосується слизької послідовності, вторинної структури РНК або обох. Сигнал -1 рибосомального зсуву рамки зчитування складається з обох елементів, розділених спейсерною областю, як правило, довжиною 5–9 нуклеотидів[2]. Зсув рамки зчитування може також бути індукований іншими молекулами, які взаємодіють з рибосомою або мРНК (транс-діючими).
Слизька послідовність
Слизькі послідовності потенційно можуть спричинити «ковзання» зчитування рибосоми та пропускання ряду нуклеотидів (зазвичай лише 1) і зчитування зовсім іншої рамки після цього. За запрограмованого -1 рибосомального зсуву рамки зчитування, слизька послідовність відповідає мотиву X_XXY_YYZ, де XXX є будь-якими трьома однаковими нуклеотидами (хоча трапляються деякі винятки), YYY це, як правило, УУУ або ААА, а Z — це А, Ц або У. У випадку з +1 зсувом рамки зчитування, слизька послідовність містить кодони, для яких відповідна тРНК є рідкісною, і зсуву рамки зчитування сприяє те, що асоційована тРНК кодону нової рамки зчитування є поширенішою[12]. Одним із прикладів слизької послідовності є полі (А) послідовність мРНК, яка, як відомо, індукує «ковзання» рибосоми навіть за відсутності будь-яких інших елементів[15].
Вторинна структура РНК
Ефективний рибосомальний зсув рамки зчитування, як правило, вимагає наявності вторинної структури РНК для посилення ефектів слизької послідовності[11]. Вважається, що структура РНК (яка може бути шпилькою або псевдовузлом) тимчасово зупиняє рибосому на слизькому сайті під час трансляції, змушуючи її переміщатись і продовжувати реплікацію з положення -1. Вважається, що це відбувається тому, що структура фізично блокує рух рибосоми, через застрягання в рибосомному мРНК тунелі[2]. Цю модель підтримує той факт, що механічна міцність псевдовузлів позитивно корелює з рівнем рибосомальних зсувів рамки зчитування для асоційованої мРНК[3][16].
Нижче наведено приклади передбачуваних вторинних структур для елементів зсувів рамки зчитування, показаних для стимулювання зсуву рамки зчитування у різних організмів. Більшість представлених структур — це шпильки, за винятком структури ALIL (англ. apical loop-internal loop) псевдовузла. На цих зображеннях більші та неповні кола мРНК представляють лінійні області. Вторинні «шпилькові» структури, де «стебла» утворені областю мРНК, основи якої спаровані з іншою областю того ж ланцюга, показані виступаючими з лінійної ДНК. Лінійна область сигналу рибосомального зсуву рамки зчитування ВІЛ містить дуже консервативну UUU UUU слизьку послідовність; багато інших прогнозованих структур містять також кандидати в слизькі послідовності.
Послідовності мРНК на зображеннях можна читати відповідно до набору вказівок. Хоча A, T, Ц і Г показують позицію певного нуклеотиду, є також букви, які показують неоднозначність і, які використовуються, коли в цій позиції може бути більше одного типу нуклеотидів. Правила Міжнародного союзу фундаментальної та прикладної хімії (IUPAC) такі[17]:
| Умовне позначення[17] | Опис | Bases represented | Complement | ||||
|---|---|---|---|---|---|---|---|
| A | Adenine | A | 1 | T | |||
| C | Cytosine | C | G | ||||
| G | Guanine | G | C | ||||
| T | Thymine | T | A | ||||
| U | Uracil | U | A | ||||
| W | Weak | A | T | 2 | W | ||
| S | Strong | C | G | S | |||
| M | aMino | A | C | K | |||
| K | Keto | G | T | M | |||
| R | puRine | A | G | Y | |||
| Y | pYrimidine | C | T | R | |||
| B | not A (B comes after A) | C | G | T | 3 | V | |
| D | not C (D comes after C) | A | G | T | H | ||
| H | not G (H comes after G) | A | C | T | D | ||
| V | not T (V comes after T and U) | A | C | G | B | ||
| N | any Nucleotide (not a gap) | A | C | G | T | 4 | N |
| Z | Zero | 0 | Z | ||||
Транс-діючі елементи
Було виявлено, що невеликі молекули, білки та нуклеїнові кислоти стимулюють рівень зсуву рамки зчитування. Наприклад, механізм негативного зворотного зв'язку в шляху синтезу поліамінів, що базується рівнях поліамінів, стимулює збільшення +1 зсувів рамок зчитування, внаслідок чого утворюється інгібіторний фермент. Було показано, що деякі білки, які необхідні для розпізнавання кодону, або які безпосередньо зв'язуються з послідовністю мРНК також модулюють рівень зсувів рамок зчитування. Молекули мікроРНК можуть гібридизуватися з вторинною структурою РНК і впливати на її міцність[5].
Див. також
Посилання
- Atkins JF, Loughran G, Bhatt PR, Firth AE, Baranov PV (вересень 2016). Ribosomal frameshifting and transcriptional slippage: From genetic steganography and cryptography to adventitious use. Nucleic Acids Research 44 (15): 7007–7078. PMC 5009743. PMID 27436286. doi:10.1093/nar/gkw530.
- Napthine S, Ling R, Finch LK, Jones JD, Bell S, Brierley I, Firth AE (червень 2017). Protein-directed ribosomal frameshifting temporally regulates gene expression. Nature Communications 8: 15582. Bibcode:2017NatCo...815582N. PMC 5472766. PMID 28593994. doi:10.1038/ncomms15582.
- Ketteler R (2012). On programmed ribosomal frameshifting: the alternative proteomes. Frontiers in Genetics (English) 3: 242. PMC 3500957. PMID 23181069. doi:10.3389/fgene.2012.00242.
- Ivanov IP, Atkins JF (2007). Ribosomal frameshifting in decoding antizyme mRNAs from yeast and protists to humans: close to 300 cases reveal remarkable diversity despite underlying conservation. Nucleic Acids Research 35 (6): 1842–1858. PMC 1874602. PMID 17332016. doi:10.1093/nar/gkm035.
- Dever TE, Dinman JD, Green R (серпень 2018). Translation Elongation and Recoding in Eukaryotes. Cold Spring Harbor Perspectives in Biology 10 (8): a032649. PMC 6071482. PMID 29610120. doi:10.1101/cshperspect.a032649.
- Jacks T, Power MD, Masiarz FR, Luciw PA, Barr PJ, Varmus HE (січень 1988). Characterization of ribosomal frameshifting in HIV-1 gag-pol expression. Nature 331 (6153): 280–283. Bibcode:1988Natur.331..280J. PMID 2447506. doi:10.1038/331280a0.
- Jacks T, Madhani HD, Masiarz FR, Varmus HE (листопад 1988). Signals for ribosomal frameshifting in the Rous sarcoma virus gag-pol region. Cell 55 (3): 447–458. PMID 2846182. doi:10.1016/0092-8674(88)90031-1.
- Jagger BW, Wise HM, Kash JC, Walters KA, Wills NM, Xiao YL, Dunfee RL, Schwartzman LM, Ozinsky A, Bell GL, Dalton RM, Lo A, Efstathiou S, Atkins JF, Firth AE, Taubenberger JK, Digard P (липень 2012). An overlapping protein-coding region in influenza A virus segment 3 modulates the host response. Science 337 (6091): 199–204. Bibcode:2012Sci...337..199J. PMC 3552242. PMID 22745253. doi:10.1126/science.1222213.
- Advani VM, Dinman JD (січень 2016). Reprogramming the genetic code: The emerging role of ribosomal frameshifting in regulating cellular gene expression. BioEssays 38 (1): 21–26. PMC 4749135. PMID 26661048. doi:10.1002/bies.201500131.
- Crick FH (серпень 1966). Codon—anticodon pairing: the wobble hypothesis. Journal of Molecular Biology 19 (2): 548–555. PMID 5969078. doi:10.1016/S0022-2836(66)80022-0.
- Brierley I (серпень 1995). Ribosomal frameshifting viral RNAs. The Journal of General Virology. 76 ( Pt 8) (8): 1885–1892. PMID 7636469. doi:10.1099/0022-1317-76-8-1885.
- An "integrated model" of programmed ribosomal frameshifting. Trends in Biochemical Sciences (English) 27 (9): 448–454. вересень 2002. PMID 12217519. doi:10.1016/S0968-0004(02)02149-7. Проігноровано невідомий параметр
|vauthors=(довідка) - Gurvich OL, Baranov PV, Gesteland RF, Atkins JF (June 2005). Expression levels influence ribosomal frameshifting at the tandem rare arginine codons AGG_AGG and AGA_AGA in Escherichia coli. Journal of Bacteriology 187 (12): 4023–4032. PMC 1151738. PMID 15937165. doi:10.1128/JB.187.12.4023-4032.2005.
- Caliskan N, Katunin VI, Belardinelli R, Peske F, Rodnina MV (червень 2014). Programmed −1 frameshifting by kinetic partitioning during impeded translocation. Cell 157 (7): 1619–1631. PMID 24949973. doi:10.1016/j.cell.2014.04.041.
- Arthur L, Pavlovic-Djuranovic S, Smith-Koutmou K, Green R, Szczesny P, Djuranovic S (липень 2015). Translational control by lysine-encoding A-rich sequences. Science Advances 1 (6): e1500154. Bibcode:2015SciA....1E0154A. PMC 4552401. PMID 26322332. doi:10.1126/sciadv.1500154.
- Hansen TM, Reihani SN, Oddershede LB, Sørensen MA (квітень 2007). Correlation between mechanical strength of messenger RNA pseudoknots and ribosomal frameshifting. Proceedings of the National Academy of Sciences of the United States of America 104 (14): 5830–5835. Bibcode:2007PNAS..104.5830H. PMC 1838403. PMID 17389398. doi:10.1073/pnas.0608668104.
- Nomenclature Committee of the International Union of Biochemistry (NC-IUB) (1984). Nomenclature for Incompletely Specified Bases in Nucleic Acid Sequences. Процитовано 4 лютого 2008.