Система винагород
Система винагород (англ. Reward System) — це група нервових структур, відповідальних за стимулювання бажань і потягів, задоволення, та іхнє позитивне підкріплення (тобто навчання). Винагорода — принадна й мотиваційна властивість стимулу, що викликає апетитивну чи консумативну поведінку. Існує поняття винагородного подразника чи стимула (англ. rewarding stimulus). Це будь-який стимул, об'єкт, подія, дія або ситуація, яка має потенціал, щоби спонукати нас до наближення й споживання того, що викличе винагороду. В системі т.зв оперантного кондиціювання (або інструментального навчання — способу вироблення умовних рефлексів, при якому на поведінку впливають за допомогою наслідків поведінки: винагороди або покарання), потенційно-винагороджувані подразники діють як позитивні підкріплювачі; і обернене твердження теж вірно: позитивні зміцнювачі — винагороджуються.
| В першу чергу винагороджуються | |
 |
 |


Первинні винагороди — це ті, які необхідні для виживання особини й її нащадків, і включають у себе:
- Гомеостатичні (приємна їжа)
- Репродуктивні (сексуальний контакт і материнство/батьківство)
А також:
- Внутрішньозумовлені винагороди — це безумовні винагороди, які є привабливими й мотивують поведінку приємними відчуттями.
- Зовнішньозумовлені винагороди (такі як гроші) — це умовні нагороди, які є привабливими й мотивуючими, але не є безумовно приємними. Їхня цінність утворена шляхом навчання. Вони можуть також стати приємними після утворення умовних рефлексів на основі внутрішньозумовлених винагород.
Визначення
У нейробіології система винагород визначається як сукупність структур головного мозку, які відповідають за винагородно-зумовлене пізнання, в тому числі позитивне підкріплення (навчання), а також бажання, симпатію і задоволення.
Анатомія системи винагороди
Мозкові структури, з яких складається система винагород, знаходяться, в першу чергу, серед «кортико-базально-ганглієво-таламічної петлі»; найбільш активна частина котрої знаходиться в базальних гангліях. Більшість шляхів, які з'єднують структури системи винагороди складаються з глутамінергічних нейронів, ГАМК-ергічних середніх шипуватих нейронів і дофамінергічних інтернейронів, хоча інші типи нейронів теж присутні. Система винагород об'єднує такі структури як: вентральну область покришки, вентральний стріатум (в першу чергу, прилегле ядро, і нюховий горбок), дорсальний стріатум (хвостате ядро і лушпину), чорна субстанцію, префронтальну кору, передню поясну кору, острівцеву (інсулярну) кору, гіпокамп, гіпоталамус, таламус (кілька ядер), субталамічне ядро, бліду кулю, парабрахіальні ядра, мигдалеподібне тіло та розширену мигдалину.
Серед шляхів, що з'єднують структури в кортико-базально-ганглієво-таламічній петлі, групи нейронів, відомі як мезолімбічні шляхи, які з'єднують вентральну тегментальну ділянку (ВТД) в прилеглому ядрі, а також пов'язаних з ними ГАМК D-1 тип середніх шипуватих нейронів у прилеглому ядрі, слугують одним з найважливіших компонентів системи винагороди, який бере пряму участь у безпосередньому сприйнятті мотиваційного компонента винагороди (тобто «хотіння»).[1] Більшість дофамінових шляхів (нейрони, що використовують нейромедіатор дофамін для передачі сигналу) відбуваються у ВТД і є частиною системи винагороди;[2] в цих шляхах допамін діє на D-1-подібні або D-2-подібні рецептори, тобто, стимулює (D-1-подібні) або інгібує (D-2-подібні) виробництво циклічного аденозинмонофосфат.[3] ГАМК-ергічні середні шипуваті нейрони в стріатумі також є компонентами системи винагороди.[4] Глутамінергічні проекції ядер в субталамічному ядрі, префронтальна кора, гіпокамп, таламус, мигдалеподібне тіло з'єднуються з іншими частинами системи винагороди через глутамінергічні шляхи.[5] Медіальний пучок переднього мозку, який складається з моноамінових нейронів, які шлють проективні волокна з декількох окремих ядер, також є частиною системи винагороди.
Центри задоволення
| Поняття | Визначення |
|---|---|
| Залежність | адаптивний стан, пов'язаний з синдромом відміни після припинення багаторазового впливу стимулу (наприклад, споживання наркотиків) |
| Сенсибілізація | посилена відповідь на стимул у результаті багаторазового впливу цього стимулу |
| Наркотична сенсибілізація або зворотна толерантність | ескалацію ефекту препарату, що виникає в результаті повторного введення в даній дозі |
| Абстинентний синдром | симптоми, які виникають після припинення регулярного вживання наркотиків |
| Фізична залежність | залежність, що включає в себе симптоми стійкої фізико-соматичної абстиненції (наприклад, втому і делірій) |
| Психологічна залежність | залежність, яка включає в себе емоційно-мотиваційні симптоми абстиненції (наприклад, дисфорію та ангедонію) |
| Підкріплюючі стимули | стимули, які збільшують ймовірність повторюваної поведінки в парі з ними |
| Винагородні стимули | стимули, які мозок інтерпретує як внутрішньозумовлені позитивні |
| Адиктивна поведінка | це поведінка, яка є одночасно винагородною і підкріплюючою |
| Наркотик | це препарат, який одночасно викликає винагородження і підкріплення |
| Розлад, викликаний вживанням психоактивних речовин | це стан, при якому використання таких речовин призводить до клінічно та функціонально значущих порушень або дистресу |
| Толерантність | зменшення ефекту лікарського засобу внаслідок повторного введення в тій самій дозі |
Задоволення є складовою частиною винагороди, але не всі нагороди приносять задоволення (наприклад, гроші не викликають задоволення, якщо ця реакція попередньо не зумовлена).[6] Подразники, які є натурально приємними, й тому привабливими, відомі як внутрішньозумовлені винагороди, тоді як подразники, які, хоч і є привабливими і мотивують «поведінку наближення», «таксис», approach behavior, але не є приємними за своєю природою, звуться зовнішньозумовленими винагородами.[7] Зовнішньозумовлені винагороди (наприклад, гроші) стають приємними в результаті асоціативного навчання на основі використання внутрішньозумовлених винагород.[8] іншими словами, зовнішні винагороди діють як мотивуючі магніти, які збурюють реакції саме «бажання», але не «задоволення» .[9] Гедонічні точки або центри задоволення (тобто, структури мозку, які опосередковують задоволення або реакції задоволення через внутрішні винагороди — в рамках системи винагороди), які були визначені станом на 15 травня 2015 року, містяться: в «равлику» прилеглого ядра, вентральній частині блідої кулі, і парабрахіальному ядрі мосту;[10][11] інсулярна, орбітофронтальна кора, ймовірно, також містять гедоністичні точки.[12]
Одночасна активація всіх гедонічних точок в межах системи винагороди вважається необхідною умовою для створення відчуття сильної ейфорії.
Кент Беридж (Kent Berridge), фахівець в області афективної нейробіології (яка вивчає невральні механізми емоцій), виявив, що солодкий («той, що сподобалося») і гіркий («той, що не сподобалося») смак продукують кожен свою специфічну орофаціальну експресію (міміку), і ці вирази були схожі у новонароджених людей, орангутанів і щурів. Це свідчить про те, що задоволення має об'єктивні особливості і є однаковим по суті у різних тварин. Більшість нейробіологічних досліджень показали, що чим більше дофаміну вивільняється у винагороду, тим більш ефективною є сама винагорода. Це називається гедонічним ефектом, який може бути змінений зусиллями, спрямованими на нагороду і, практично, винагороджувати себе. Але Берридж (Berridge) виявив, що блокування дофамінових систем не змінює позитивної реакції на солодощі, виміряної за виразом обличчя. Іншими словами, гедоністичний вплив не змінюються в залежності від кількості цукру. Це суперечить загальноприйнятому припущенню, що дофамін опосередковує задоволення. Навіть з більш інтенсивним коливанням дофаміну, кінцевий результат, як виявляється, залишаються незмінними. Берридж розробив гіпотезу значущості стимулу, що описує аспект очікування винагороди. Це пояснює і компульсивне вживання наркотиків наркоманами, навіть коли препарат вже не викликає ейфорію, і потяг до препаратів, котрий відчувається навіть після того, як людина успішно пролікована. Деякі залежні реагують на певні невральні зміни, викликані наркотиками. Ця сенсибілізація мозкових структур схожа на ефект допаміну, бо в цьому випадку також виникають реакції бажання та задоволення. Мозок як людини, так і тварин переживає аналогічні зміни, що стосуються системи винагороди через подібність процесів.
Історія
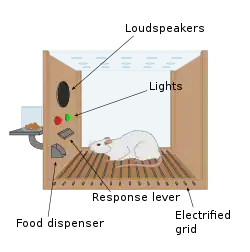
Джеймс Олдс(James Olds) і Пітер Мілнер (Peter Milner) були дослідниками, які відкрили систему винагород в 1954 році, намагаючись навчити щурів як вирішувати проблеми і проходити лабіринти. Вони подразнювали певні ділянки мозку, стимуляція котрих, як було згодом виявлено, викликає задоволення у тварин. Вони спробували провести такі самі досліди з людьми й результати були схожими.
У фундаментальному відкритті, зробленім у 1954 році, дослідники Джеймс Олдс і Пітер Мілнер виявили, що електрична стимуляція певних ділянок мозку струмами низького вольтажу у щурів може слугувати винагородою при навчанні тварин проходити лабіринти і вирішувати проблеми. Виявилося, що стимуляція цих частин мозку давали тваринам задоволення і в більш пізній роботі також люди повідомляли про приємні відчуття від такої стимуляції. Коли щури були протестовані в боксі Скінера (англ. Skinner boxes), де була можливість самостійно стимулювати ділянки системи винагород шляхом натискання на важіль, вони тиснули його протягом кількох годин. Дослідженнями протягом наступних двох десятиліть встановлено, що дофамін є однією з основних хімічних речовин, які допомагають проходженню нейронних сигналів в цих ділянках, і запропоновано вважати його «хімічним задоволенням мозку».
Іван Павлов був психологом, який використовував систему винагород для експериментального створення умовних рефлексів. Павлов використовував систему винагороди практично — шляхом заохочення собак їжею після того, як вони почули дзвінок або інший стимул. Павлов винагороджував собак так, що вони пов'язували нагороду за їжу з дзвінком чи іншим стимулом.
Едвард Л. Торндайк (Edward L. Thorndike) використовував системи винагороди для вивчення оперантного кондиціонування. Він помістив кішку в коробку з головоломкою, а їжу розміщував поза коробкою так, щоб кішка хотіла втекти до їжі. Торндайк побачив, що кішки тікали з коробки, коли була їжа, також вони намагалися втекти без нагороди їжею. Торндайк давав котам їжу і свободу для стимуляції системи винагороди, щоби побачити, як кішки вчаться тікати з коробки.
Залежність
ΔFosB (delta FosB), фактор транскрипції генів, є спільним фактором для практично всіх форм залежності (як поведінкової так і наркотичної). Коли концентрація його перевищена в D1-типу середніх шипуватих нейронах у прилеглому ядрі, то виникає поведінкова залежність та залежність нейронної пластичності, зокрема, ΔFosB сприяє «самопризначенню» препарату (рішенню про його вживання), підвищенню «винагородної сенсибілізації» (процесу, який викликає збільшення кількості винагороди) і перехресної «винагородної сенсибілізації» між конкретними наркотиками і поведінковими залежностями.
Наркотики і адиктивна поведінка винагороджуються і підкріплюються (тобто, виникає звикання) у зв'язку з їх впливом на дофаміновий мезолімбічний шлях. Вони не тільки стимулюють ці мозкові системи винагороди більш енергійно, ніж природні нагороди, але можуть також змінити їх. Постійне вживання наркотиків та участь у діяльності, що викликає залежність, знижує резистентність і спричиняє виникнення толерантності. У той час як різке припинення вживання препарату й припинення адиктивної діяльності викликає падіння концентрації дофаміну нижче нормального рівня і спричиняє абстинентний синдром.
Див. також
Примітки
- Nestler EJ (December 2013)"Reward and the NAcc shell vs core", authors: Dumitriu D, Laplant Q, Grossman YS, Dias C, Janssen WG, Russo SJ, Morrison JH, Nestler Тitle: Subregional, dendritic compartment, and spine subtype specificity in cocaine regulation of dendritic spines in the nucleus accumbens. J. Neurosci.volume32, issue20, pages — 6957–66, year-2012. quote: The enduring spine density change in core but not shell fits well with the established idea that the shell is preferentially involved in the development of addiction, while the core mediates the long-term execution of learned addiction-related behaviors (Ito et al., 2004; Di Chiara, 2002; Meredith et al., 2008). Consistent with the idea of NAc core being the locus of long-lasting drug-induced neuroplasticity, several studies have shown that electrophysiological changes in core persist longer than their shell counterparts. … Furthermore, data presented here support the idea that NAc shell is preferentially involved in immediate drug reward, while the core might play a more explicit role in longer-term aspects of addiction.
- The ins and outs of the striatum: Role in drug addiction, journal Neuroscience «Striatal efferents, afferents, and colocalized receptors in dMSNs and iMSNs», authors: Yager LM, Garcia AF, Wunsch AM, Ferguson SM volume-301, pages 529—541, date-August 2015 quote: The [striatum] receives dopaminergic inputs from the ventral tegmental area (VTA) and the substantia nigra (SNr) and glutamatergic inputs from several areas, including the cortex, hippocampus, amygdala, and thalamus (Swanson, 1982; Phillipson and Griffiths, 1985; Finch, 1996; Groenewegen et al., 1999; Britt et al., 2012). These glutamatergic inputs make contact on the heads of dendritic spines of the striatal GABAergic medium spiny projection neurons (MSNs) whereas dopaminergic inputs synapse onto the spine neck, allowing for an important and complex interaction between these two inputs in modulation of MSN activity … It should also be noted that there is a small population of neurons in the NAc that coexpress both D1 and D2 receptors, though this is largely restricted to the NAc shell (Bertran- Gonzalez et al., 2008). … Neurons in the NAc core and NAc shell subdivisions also differ functionally. The NAc core is involved in the processing of conditioned stimuli whereas the NAc shell is more important in the processing of unconditioned stimuli; Classically, these two striatal MSN populations are thought to have opposing effects on basal ganglia output. Activation of the dMSNs causes a net excitation of the thalamus resulting in a positive cortical feedback loop; thereby acting as a ‘go’ signal to initiate behavior. Activation of the iMSNs, however, causes a net inhibition of thalamic activity resulting in a negative cortical feedback loop and therefore serves as a ‘brake’ to inhibit behavior … there is also mounting evidence that iMSNs play a role in motivation and addiction (Lobo and Nestler, 2011; Grueter et al., 2013). For example, optogenetic activation of NAc core and shell iMSNs suppressed the development of a cocaine CPP whereas selective ablation of NAc core and shell iMSNs … enhanced the development and the persistence of an amphetamine CPP (Durieux et al., 2009; Lobo et al., 2010). These findings suggest that iMSNs can bidirectionally modulate drug reward. … Together these data suggest that iMSNs normally act to restrain drug-taking behavior and recruitment of these neurons may in fact be protective against the development of compulsive drug use.
- Trantham-Davidson H., Neely L. C., Lavin A., Seamans J. K. Mechanisms underlying differential D1 versus D2 dopamine receptor regulation of inhibition in prefrontal cortex. The Journal of Neuroscience, volume24, issue=47, pages=10652–10659
- journal Neuroscience, volume 301, pages529–541, August 2015 «The ins and outs of the striatum». Yager LM, Garcia AF, Wunsch AM, Ferguson SM. Striatal efferents, afferents, and colocalized receptors in dMSNs and iMSNs. : Role in drug addiction. quote: [The striatum] receives dopaminergic inputs from the ventral tegmental area (VTA) and the substantia nigra (SNr) and glutamatergic inputs from several areas, including the cortex, hippocampus, amygdala, and thalamus (Swanson, 1982; Phillipson and Griffiths, 1985; Finch, 1996; Groenewegen et al., 1999; Britt et al., 2012). These glutamatergic inputs make contact on the heads of dendritic spines of the striatal GABAergic medium spiny projection neurons (MSNs) whereas dopaminergic inputs synapse onto the spine neck, allowing for an important and complex interaction between these two inputs in modulation of MSN activity … It should also be noted that there is a small population of neurons in the NAc that coexpress both D1 and D2 receptors, though this is largely restricted to the NAc shell (Bertran- Gonzalez et al., 2008). … Neurons in the NAc core and NAc shell subdivisions also differ functionally. The NAc core is involved in the processing of conditioned stimuli whereas the NAc shell is more important in the processing of unconditioned stimuli; Classically, these two striatal MSN populations are thought to have opposing effects on basal ganglia output. Activation of the dMSNs causes a net excitation of the thalamus resulting in a positive cortical feedback loop; thereby acting as a ‘go’ signal to initiate behavior. Activation of the iMSNs, however, causes a net inhibition of thalamic activity resulting in a negative cortical feedback loop and therefore serves as a ‘brake’ to inhibit behavior … there is also mounting evidence that iMSNs play a role in motivation and addiction (Lobo and Nestler, 2011; Grueter et al., 2013). For example, optogenetic activation of NAc core and shell iMSNs suppressed the development of a cocaine CPP whereas selective ablation of NAc core and shell iMSNs … enhanced the development and the persistence of an amphetamine CPP (Durieux et al., 2009; Lobo et al., 2010). These findings suggest that iMSNs can bidirectionally modulate drug reward. … Together these data suggest that iMSNs normally act to restrain drug-taking behavior and recruitment of these neurons may in fact be protective against the development of compulsive drug use.
- Striatal efferents, afferents, and colocalized receptors in dMSNs and iMSNs" Authors: Yager LM, Garcia AF, Wunsch AM, Ferguson SM Title: The ins and outs of the striatum: Role in drug addiction. journal Neuroscience, volume=301, pages=529–541, August 2015. Quote: [The striatum] receives dopaminergic inputs from the ventral tegmental area (VTA) and the substantia nigra (SNr) and glutamatergic inputs from several areas, including the cortex, hippocampus, amygdala, and thalamus (Swanson, 1982; Phillipson and Griffiths, 1985; Finch, 1996; Groenewegen et al., 1999; Britt et al., 2012). These glutamatergic inputs make contact on the heads of dendritic spines of the striatal GABAergic medium spiny projection neurons (MSNs) whereas dopaminergic inputs synapse onto the spine neck, allowing for an important and complex interaction between these two inputs in modulation of MSN activity … It should also be noted that there is a small population of neurons in the NAc that coexpress both D1 and D2 receptors, though this is largely restricted to the NAc shell (Bertran- Gonzalez et al., 2008). … Neurons in the NAc core and NAc shell subdivisions also differ functionally. The NAc core is involved in the processing of conditioned stimuli whereas the NAc shell is more important in the processing of unconditioned stimuli; Classically, these two striatal MSN populations are thought to have opposing effects on basal ganglia output. Activation of the dMSNs causes a net excitation of the thalamus resulting in a positive cortical feedback loop; thereby acting as a ‘go’ signal to initiate behavior. Activation of the iMSNs, however, causes a net inhibition of thalamic activity resulting in a negative cortical feedback loop and therefore serves as a ‘brake’ to inhibit behavior … there is also mounting evidence that iMSNs play a role in motivation and addiction (Lobo and Nestler, 2011; Grueter et al., 2013). For example, optogenetic activation of NAc core and shell iMSNs suppressed the development of a cocaine CPP whereas selective ablation of NAc core and shell iMSNs … enhanced the development and the persistence of an amphetamine CPP (Durieux et al., 2009; Lobo et al., 2010). These findings suggest that iMSNs can bidirectionally modulate drug reward. … Together these data suggest that iMSNs normally act to restrain drug-taking behavior and recruitment of these neurons may in fact be protective against the development of compulsive drug use.
- Schultz W, 2015 Neuronal reward and decision signals: from theories to data.Journal Physiological Reviews, volume95, issue3, pages 853—951 archivedate 6 September 2015 Quote: Rewards in operant conditioning are positive reinforcers. … Operant behavior gives a good definition for rewards. Anything that makes an individual come back for more is a positive reinforcer and therefore a reward. Although it provides a good definition, positive reinforcement is only one of several reward functions. … Rewards are attractive. They are motivating and make us exert an effort. … Rewards induce approach behavior, also called appetitive or preparatory behavior, and consummatory behavior. … Thus any stimulus, object, event, activity, or situation that has the potential to make us approach and consume it is by definition a reward. … Rewarding stimuli, objects, events, situations, and activities consist of several major components. First, rewards have basic sensory components (visual, auditory, somatosensory, gustatory, and olfactory) … Second, rewards are salient and thus elicit attention, which are manifested as orienting responses (FIGURE 1, middle). The salience of rewards derives from three principal factors, namely, their physical intensity and impact (physical salience), their novelty and surprise (novelty/surprise salience), and their general motivational impact shared with punishers (motivational salience). A separate form not included in this scheme, incentive salience, primarily addresses dopamine function in addiction and refers only to approach behavior (as opposed to learning) … Third, rewards have a value component that determines the positively motivating effects of rewards and is not contained in, nor explained by, the sensory and attentional components (FIGURE 1, right). This component reflects behavioral preferences and thus is subjective and only partially determined by physical parameters. Only this component constitutes what we understand as a reward. It mediates the specific behavioral reinforcing, approach generating, and emotional effects of rewards that are crucial for the organism's survival and reproduction, whereas all other components are only supportive of these functions. … Rewards can also be intrinsic to behavior (31, 546, 547). They contrast with extrinsic rewards that provide motivation for behavior and constitute the essence of operant behavior in laboratory tests. Intrinsic rewards are activities that are pleasurable on their own and are undertaken for their own sake, without being the means for getting extrinsic rewards. … Intrinsic rewards are genuine rewards in their own right, as they induce learning, approach, and pleasure, like perfectioning, playing, and enjoying the piano. Although they can serve to condition higher order rewards, they are not conditioned, higher order rewards, as attaining their reward properties does not require pairing with an unconditioned reward. … These emotions are also called liking (for pleasure) and wanting (for desire) in addiction research (471) and strongly support the learning and approach generating functions of reward.
- Schultz W, 2015, Neuronal reward and decision signals: from theories to data, journal Physiological Reviews, volume=95, issue 3, pages 853—951, , archive Quote Rewards in operant conditioning are positive reinforcers. … Operant behavior gives a good definition for rewards. Anything that makes an individual come back for more is a positive reinforcer and therefore a reward. Although it provides a good definition, positive reinforcement is only one of several reward functions. … Rewards are attractive. They are motivating and make us exert an effort. … Rewards induce approach behavior, also called appetitive or preparatory behavior, and consummatory behavior. … Thus any stimulus, object, event, activity, or situation that has the potential to make us approach and consume it is by definition a reward. … Rewarding stimuli, objects, events, situations, and activities consist of several major components. First, rewards have basic sensory components (visual, auditory, somatosensory, gustatory, and olfactory) … Second, rewards are salient and thus elicit attention, which are manifested as orienting responses (FIGURE 1, middle). The salience of rewards derives from three principal factors, namely, their physical intensity and impact (physical salience), their novelty and surprise (novelty/surprise salience), and their general motivational impact shared with punishers (motivational salience). A separate form not included in this scheme, incentive salience, primarily addresses dopamine function in addiction and refers only to approach behavior (as opposed to learning) … Third, rewards have a value component that determines the positively motivating effects of rewards and is not contained in, nor explained by, the sensory and attentional components (FIGURE 1, right). This component reflects behavioral preferences and thus is subjective and only partially determined by physical parameters. Only this component constitutes what we understand as a reward. It mediates the specific behavioral reinforcing, approach generating, and emotional effects of rewards that are crucial for the organism's survival and reproduction, whereas all other components are only supportive of these functions. … Rewards can also be intrinsic to behavior (31, 546, 547). They contrast with extrinsic rewards that provide motivation for behavior and constitute the essence of operant behavior in laboratory tests. Intrinsic rewards are activities that are pleasurable on their own and are undertaken for their own sake, without being the means for getting extrinsic rewards. … Intrinsic rewards are genuine rewards in their own right, as they induce learning, approach, and pleasure, like perfectioning, playing, and enjoying the piano. Although they can serve to condition higher order rewards, they are not conditioned, higher order rewards, as attaining their reward properties does not require pairing with an unconditioned reward. … These emotions are also called liking (for pleasure) and wanting (for desire) in addiction research (471) and strongly support the learning and approach generating functions of reward.
- Schultz W, 2015, Neuronal reward and decision signals: from theories to data, journal Physiological Reviews, volume-95, issue3, pages 853—951 archive Quote Rewards in operant conditioning are positive reinforcers. … Operant behavior gives a good definition for rewards. Anything that makes an individual come back for more is a positive reinforcer and therefore a reward. Although it provides a good definition, positive reinforcement is only one of several reward functions. … Rewards are attractive. They are motivating and make us exert an effort. … Rewards induce approach behavior, also called appetitive or preparatory behavior, and consummatory behavior. … Thus any stimulus, object, event, activity, or situation that has the potential to make us approach and consume it is by definition a reward. … Rewarding stimuli, objects, events, situations, and activities consist of several major components. First, rewards have basic sensory components (visual, auditory, somatosensory, gustatory, and olfactory) … Second, rewards are salient and thus elicit attention, which are manifested as orienting responses (FIGURE 1, middle). The salience of rewards derives from three principal factors, namely, their physical intensity and impact (physical salience), their novelty and surprise (novelty/surprise salience), and their general motivational impact shared with punishers (motivational salience). A separate form not included in this scheme, incentive salience, primarily addresses dopamine function in addiction and refers only to approach behavior (as opposed to learning) … Third, rewards have a value component that determines the positively motivating effects of rewards and is not contained in, nor explained by, the sensory and attentional components (FIGURE 1, right). This component reflects behavioral preferences and thus is subjective and only partially determined by physical parameters. Only this component constitutes what we understand as a reward. It mediates the specific behavioral reinforcing, approach generating, and emotional effects of rewards that are crucial for the organism's survival and reproduction, whereas all other components are only supportive of these functions. … Rewards can also be intrinsic to behavior (31, 546, 547). They contrast with extrinsic rewards that provide motivation for behavior and constitute the essence of operant behavior in laboratory tests. Intrinsic rewards are activities that are pleasurable on their own and are undertaken for their own sake, without being the means for getting extrinsic rewards. … Intrinsic rewards are genuine rewards in their own right, as they induce learning, approach, and pleasure, like perfectioning, playing, and enjoying the piano. Although they can serve to condition higher order rewards, they are not conditioned, higher order rewards, as attaining their reward properties does not require pairing with an unconditioned reward. … These emotions are also called liking (for pleasure) and wanting (for desire) in addiction research (471) and strongly support the learning and approach generating functions of reward.
- Schultz W, 2015, Neuronal reward and decision signals: from theories to data, Physiological Reviews, v95,i3, pages 853—951 Quote Rewards in operant conditioning are positive reinforcers. … Operant behavior gives a good definition for rewards. Anything that makes an individual come back for more is a positive reinforcer and therefore a reward. Although it provides a good definition, positive reinforcement is only one of several reward functions. … Rewards are attractive. They are motivating and make us exert an effort. … Rewards induce approach behavior, also called appetitive or preparatory behavior, and consummatory behavior. … Thus any stimulus, object, event, activity, or situation that has the potential to make us approach and consume it is by definition a reward. … Rewarding stimuli, objects, events, situations, and activities consist of several major components. First, rewards have basic sensory components (visual, auditory, somatosensory, gustatory, and olfactory) … Second, rewards are salient and thus elicit attention, which are manifested as orienting responses (FIGURE 1, middle). The salience of rewards derives from three principal factors, namely, their physical intensity and impact (physical salience), their novelty and surprise (novelty/surprise salience), and their general motivational impact shared with punishers (motivational salience). A separate form not included in this scheme, incentive salience, primarily addresses dopamine function in addiction and refers only to approach behavior (as opposed to learning) … Third, rewards have a value component that determines the positively motivating effects of rewards and is not contained in, nor explained by, the sensory and attentional components (FIGURE 1, right). This component reflects behavioral preferences and thus is subjective and only partially determined by physical parameters. Only this component constitutes what we understand as a reward. It mediates the specific behavioral reinforcing, approach generating, and emotional effects of rewards that are crucial for the organism's survival and reproduction, whereas all other components are only supportive of these functions. … Rewards can also be intrinsic to behavior (31, 546, 547). They contrast with extrinsic rewards that provide motivation for behavior and constitute the essence of operant behavior in laboratory tests. Intrinsic rewards are activities that are pleasurable on their own and are undertaken for their own sake, without being the means for getting extrinsic rewards. … Intrinsic rewards are genuine rewards in their own right, as they induce learning, approach, and pleasure, like perfectioning, playing, and enjoying the piano. Although they can serve to condition higher order rewards, they are not conditioned, higher order rewards, as attaining their reward properties does not require pairing with an unconditioned reward. … These emotions are also called liking (for pleasure) and wanting (for desire) in addiction research (471) and strongly support the learning and approach generating functions of reward.
- Berridge KC, Kringelbach ML, Pleasure systems in the brain, journal Neuron v86 issue3 pages 646—664,May 2015 Quote: In the prefrontal cortex, recent evidence indicates that the OFC and insula cortex may each contain their own additional hot spots (D.C. Castro et al., Soc. Neurosci., abstract). In specific subregions of each area, either opioid-stimulating or orexin-stimulating microinjections appear to enhance the number of ‘‘liking’’ reactions elicited by sweetness, similar to the NAc and VP hot spots. Successful confirmation of hedonic hot spots in the OFC or insula would be important and possibly relevant to the orbitofrontal mid-anterior site mentioned earlier that especially tracks the subjective pleasure of foods in humans (Georgiadis et al., 2012; Kringelbach, 2005; Kringelbach et al., 2003; Small et al., 2001; Veldhuizen et al., 2010). Finally, in the brainstem, a hindbrain site near the parabrachial nucleus of dorsal pons also appears able to contribute to hedonic gains of function (Söderpalm and Berridge, 2000). A brainstem mechanism for pleasure may seem more surprising than forebrain hot spots to anyone who views the brainstem as merely reflexive, but the pontine parabrachial nucleus contributes to taste, pain, and many visceral sensations from the body and has also been suggested to play an important role in motivation (Wu et al., 2012) and in human emotion (especially related to the somatic marker hypothesis) (Damasio, 2010).
- Richard JM, Castro DC, Difeliceantonio AG, Robinson MJ, Berridge KC, Mapping brain circuits of reward and motivation: in the footsteps of Ann Kelley, journal Neurosci. Biobehav. Rev. volume37 issue9 Pt A pages 1919—1931, November 2013 Quote Figure 3: Neural circuits underlying motivated 'wanting' and hedonic 'liking'.
- Berridge KC, Kringelbach ML: Pleasure systems in the brain, journal Neuron, volume-86, issue3, pages-646–664, May 2015 Quote=In the prefrontal cortex, recent evidence indicates that the OFC and insula cortex may each contain their own additional hot spots (D.C. Castro et al., Soc. Neurosci., abstract). In specific subregions of each area, either opioid-stimulating or orexin-stimulating microinjections appear to enhance the number of ‘‘liking’’ reactions elicited by sweetness, similar to the NAc and VP hot spots. Successful confirmation of hedonic hot spots in the OFC or insula would be important and possibly relevant to the orbitofrontal mid-anterior site mentioned earlier that especially tracks the subjective pleasure of foods in humans (Georgiadis et al., 2012; Kringelbach, 2005; Kringelbach et al., 2003; Small et al., 2001; Veldhuizen et al., 2010). Finally, in the brainstem, a hindbrain site near the parabrachial nucleus of dorsal pons also appears able to contribute to hedonic gains of function (Söderpalm and Berridge, 2000). A brainstem mechanism for pleasure may seem more surprising than forebrain hot spots to anyone who views the brainstem as merely reflexive, but the pontine parabrachial nucleus contributes to taste, pain, and many visceral sensations from the body and has also been suggested to play an important role in motivation (Wu et al., 2012) and in human emotion (especially related to the somatic marker hypothesis) (Damasio, 2010).
Посилання
- Мозкові шляхи — Опис системи винагороди, опубліковані Nestler Лабораторія, Кафедра неврології, школи медицини Ікан
- Scholarpedia: Винагорода
- Scholarpedia Сигнали системи винагороди