Вірус гепатиту D
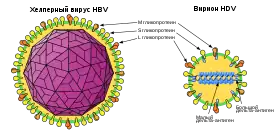
Вірус гепати́ту дельта, або вірус гепати́ту D (HDV)— інфекційний агент, що спричинює гепатит D у людини. Цей невеликий РНК-вмісний інфекційний агент є вірусом-сателітом, позаяк для його розмноження в клітинах і розвитку інфекції необхідно, щоб клітини були заражені вірусом гепатиту B (HBV). HDV використовує оболонкові білки вірусу гепатиту B (HBsAg) для упакування свого геному.
? Вірус гепатиту D | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||
| Класифікація вірусів | ||||||||||
| ||||||||||
Посилання | ||||||||||
| ||||||||||
Вірус гепатиту дельта спочатку був описаний у пацієнтів із важкою формою інфекції, що спричинюється вірусом гепатиту B. Зараження гепатитом D може відбуватись і разом із зараженням гепатитом B (коінфекція), і накладатися на хронічний гепатит B (суперінфекція). В обох випадках у пацієнтів виявляються важчі симптоми порівняно з самим лише гепатитом B. Серед них набагато вищою є ймовірність розвитку кінцевої стадії печінкової недостатності внаслідок гострої інфекції, швидкого розвитку цирозу печінки, а у випадку хронічних інфекцій — підвищена ймовірність печінковоклітинної карциноми.
Вірус гепатиту дельта є винятковим серед патогенів людини та тварин через те, що він має низку спільних властивостей із віроїдами рослин і з віроїд-подібними сателітними РНК рослин. Цей патоген, що передається кров'ю, розмножується в печінці та може спричинювати гострий гепатит як у приматів, так і ссавців, що не належать до приматів (хоча єдиним господарем вірусу є людина). У світі вірусом гепатиту дельта заражені понад 15 мільйонів людей, що робить його важливою проблемою сучасної охорони здоров'я.
Історія вивчення
Перші повідомлення про вірус гепатиту дельта з'явились у середині 1977 року. Його відкрив Маріо Ріцетто з колегами, разом вони вивчали групу пацієнтів, що були інфіковані вірусом гепатиту B та страждали від особливо гострої форми гепатиту. Він був описаний як новий ядерний антиген вірусу гепатиту B і одержав назву антиген дельта (δ, HDAg). Подальші експерименти на шимпанзе показали, що дельта-антиген насправді є структурним елементом патогену, для реплікації якого був необхідний вірус гепатиту B. До 1980 року вірус гепатиту дельта не вважали інфекційним агентом. Однак незабаром після визнання вірусу гепатиту D патогеном були розроблені ефективні тести на його виявлення. Окрім цього було відкрито збір епідеміологічної інформації щодо гепатиту D (він розпочався з Італії). Геном вірусу гепатиту дельта клонували та секвенували 1986 року. 1993 року вірус зареєстрував Міжнародний комітет із таксономії вірусів і помістив його до монотипного роду Deltavirus.
Еволюція та походження
Природним господарем HDV є людина. Дані філогенетичних досліджень вказують на африканське походження вірусу гепатиту дельта. HDV характеризується високим ступенем генетичної гетерогенності. Вважається, що еволюцію HDV забезпечують 3 основних механізми: мутації, редагування та рекомбінація. Швидкість мутування становить, за різними оцінками, від от 3·10−2 до 3·10−3 замін на геном щороку. Вона залежить від фази інфекції (є найвищою в гострій фазі), ділянки геному (висока в неконсервативних ділянках і низька в консервативних, наприклад, у області рибозиму) та зростає від терапевтичного тиску. Швидкість мутування HDV є вищою, ніж у більшості РНК-вмісних вірусів. У зв'язку з такою швидкістю мутування припускається, що HDV циркулює в межах одного зараженого організму-господаря як низка квазівидів. Встановлено, що до 70 % замін можуть бути обумовлені редагуванням. Уперше рекомбінацію в HDV описали 1999 року; тоді було зроблено висновок, що вона не відбувається у випадку зараження вірусами різних генотипів. Рекомбінація відбувається гомологічним шляхом. Припускається, що в рекомбінації в HDV бере участь РНК-полімераза клітини-господаря.
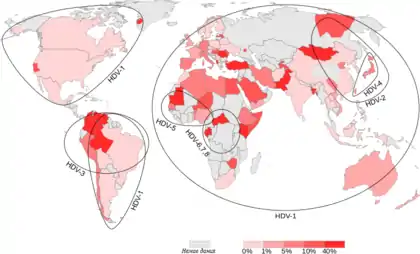
Спершу було описано 3 генотипи цього вірусу (I—III). Генотип I було виділено в Європі, Північній Америці, Африці та деяких реґіонах Азії. Генотип II спостерігають у Японії, на Тайвані та в Якутії. Генотип III виявляють лише в Південній Америці (Перу, Колумбія та Венесуела). Зараз відомо, що існує щонайменше 8 генотипів вірусу гепатиту дельта (HDV-1 — HDV-8). Усі вони, за винятком HDV-1, прив'язані до строго визначених географічних реґіонів. HDV-2 (раніше відомий як HDV-IIa) знайдений у Японії, на Тайвані та в Якутії; HDV-4 (HDV-IIb) — в Японії та на Тайвані; HDV-3 — в районі Амазонки; HDV-5, HDV-6, HDV-7 і HDV-8 — в Африці.
Сьогодні поширені дві основні теорії щодо походження вірусу гепатиту дельта. Відповідно до них, HDV походить від віроїдів рослин і/або в наслідок сплайсинґу пре-мРНК клітини-господаря. РНК HDV за особливостями структури та реплікації має спільні риси з кожною з двох родин віроїдів, відомих станом на сьогодні (Pospiviroidae й Avsunviroidae). З Pospiviroidae цей вірус поєднує паличкоподібна структура РНК та реплікація в ядрі, а з Avsunviroidae — наявність рибозиму та симетрична реплікація за типом рухомого кола. Окрім того, РНК HDV та віроїдів рослин взаємодіють із гомологічними клітинними білками, а експериментальні дані 2012 року показують, що HDV може реплікуватись і розмножуватись після його внесення в листки сходів помідора, що слугує ще одним підтвердженням близькості HDV та віроїдів. Однак ця гіпотеза не відповідає на питання щодо походження дельта-антигену та зв'язку HDV з HBV.
Друга теорія, яка може доповнювати першу, полягає в тому, що HDV може походити з транскриптому клітини-господаря. Ця точка зору підтверджується дослідженнями, що показали, що в клітинах людини є рибозим (в інтроні гену CPEB3), за вторинною структурою та біохімічними властивостями схожий із рибозимом HDV. Втім рибозими, що мають структурний елемент псевдовузол, пізніше були виявлені у всіх царствах живих організмів, за винятком археїв, а також у вірусах комах. Для дельта-антигену також припускається походження з клітини-господаря. Спершу можливим білком-предком дельта-антигену вважали білок DIPA (англ. delta interacting protein A.Хоча зрештою виявилось, що ці білки не гомологічні, DIPA може взаємодіяти з HDAg.
Інтегрована модель припускає, що HDV міг виникнути після рекомбінації між віроїд-подібним елементом і клітинною пре-мРНК/мРНК.
Література
- Fields Virology / Editors-in-chief David M. Knipe, Peter M. Howley. — Sixth edition. — Philadelphia, USA : Lippincott Williams & Wilkins, 2013. — 2582 p. — ISBN 978-1-4511-0563-6.
- Nicholas H. Acheson. Fundamentals of Molecular Virology. — 2nd edition. — WILEY (John Wiley & Sons, Inc.), 2011. — P. 379—383. — ISBN 978-0-470-90059-8.
- Desk Encyclopedia of Human and Medical Virology / Editors-in-chief Brian W. J. Mahy, Marc H. V. van Regenmortel. — Elsevier Ltd, 2010. — 661 p. — ISBN 978-0-12-375147-8.
