Еволюція вірусів
Еволюція вірусів — зміни спадкових характеристик вірусів у поколіннях, а також галузь еволюційної біології та вірусології, що їх вивчає[1][2]. Віруси мають короткий період розмноження, а деякі (зокрема РНК-віруси) мають підвищену частоту мутацій (одна або більше точкових мутацій на геном за раунд реплікації). Підвищена частота мутацій разом із природним відбором дають змогу вірусам швидко адаптуватись до змін у їхньому середовищі існування. При цьому, більшість вірусів створюють численне потомство, внаслідок чого будь-який мутований ген швидко передається великій кількості нащадків. Хоч ймовірність мутації та еволюції варіює залежно від типу вірусу (дволанцюгова ДНК, дволанцюгова РНК, одноланцюгова ДНК тощо), для вірусів характерна висока частота мутацій.
Еволюція вірусів є важливим аспектом епідеміології вірусних хвороб на кшталт грипу (ортоміксовіруси), ВІЛ-інфекції (вірус імунодефіциту людини) та вірусних гепатитів (наприклад, гепатиту C, який спричинює відповідний вірус). Швидкість мутацій може стати на заваді розробки успішних вакцин та противірусних ліків, оскільки часто через декілька тижнів або місяців після початку лікування виникають мутації опірності. Однією з основних теоретичних моделей, що застосовуються до еволюції вірусів, є модель квазівидів, яка визначає квазівиди вірусів групою дуже близьких вірусних штамів, що борються за середовище.
Походження вірусів
Віруси є стародавніми організмами. Дослідження на молекулярному рівні з'ясували взаємозв'язки між вірусами, що інфікують організми з кожного із трьох доменів живого, а також передбачили вірусні білки, які передували дивергенції живих організмів, а тому заразили останнього універсального спільного предка[3]. Такі дані вказують на те, що віруси виникли на ранніх етапах еволюції,[4] і що вони, ймовірно, з'являлись декілька разів[5]. Наводились припущення, що на всіх етапах еволюції щоразу виникали нові групи вірусів, часто завдяки зміщенню стародавніх структурних і геномореплікувальних генів.[6]
Існує три класичні гіпотези щодо походження та еволюції вірусів:
- Гіпотеза першості вірусів: Віруси еволюціювали з комплексних молекул білків і нуклеїнових кислот ще до першої появи клітин на Землі.[1][2] Згідно з гіпотезою першості вірусів, віруси доклались до появи клітинного життя.[7] Ця гіпотеза підтверджується ідеєю про те, що всі вірусні геноми кодують білки, які не мають клітинних гомологів. Гіпотеза першості вірусів відкидається деякими науковцями, оскільки вона суперечить визначенню вірусів, а саме, що віруси потребують клітину-господаря для реплікації.
- Редукційна гіпотеза (гіпотеза виродження): Віруси колись були дрібними клітинами, які паразитували в більших клітинах.[5][8] Така гіпотеза підтверджується відкриттям велетенських вірусів зі схожим генетичним матеріалом із паразитичними бактеріями. Утім, редукційна гіпотеза не пояснює, чому навіть найменші клітинні паразити навіть найменшим чином не мають схожості із вірусами.[7]
- Гіпотеза випадіння (гіпотеза блукаючих генів): Деякі віруси еволюціонували з ділянок ДНК або РНК, які «випали» з генів більших організмів.[9] Ця гіпотеза не пояснює структур, що є унікальними для вірусів, і не мають ніяких проявів у клітинах. Окрім цього, гіпотеза випадіння не пояснює наявність комплексних капсидів та інших структур вірусних частинок.[7]
Утім, вірусологи в процесі оцінювання наново цих гіпотез.[6][10][11]
Пізніші гіпотези
- Гіпотеза коеволюції (теорія бульбашок): На початку життя спільнота ранніх репліконів (фрагментів генетичної інформації, здатних до самовідтворення) існувала поблизу джерела їжі, такого як термальні води або гідротермальні джерела. Це джерело їжі також виробляло ліпідоподібні молекули, які самостійно збиралися у везикули, які могли помістити реплікони. Реплікони процвітали близько до джерела їжі, але далі єдині нерозбавлені ресурси були б всередині везикул. Тому еволюційний тиск може штовхати репліканів двома шляхами розвитку: вони зливаються з везикулою, даючи початок клітинам; і потрапляючи у везикулу, використовують її ресурси, розмножуються і відправляються в іншу везикулу, утворюючи віруси.[12]
- Гіпотеза про химерне походження: На основі аналізу еволюції реплікативних та структурних модулів вірусів у 2019 році було запропоновано химерний сценарій походження вірусів. Згідно з цією гіпотезою, реплікаційні модулі вірусів походять з первинного генетичного пулу, хоча тривалий шлях їх подальшої еволюції передбачав багато переміщень реплікативних генів з їх клітинних господарів. На відміну від них, гени, що кодують основні структурні білки, еволюціонували з функціонально різноманітних білків-господарів протягом усієї еволюції віросфери. Цей сценарій відрізняється від кожного з трьох традиційних сценаріїв, але поєднує в собі особливості гіпотези про віруси та втечу.
Однією з проблем вивчення походження та еволюції вірусів є високий рівень вірусних мутацій, особливо у випадку з РНК-ретровірусами, такими як ВІЛ / СНІД. Нещодавнє дослідження, яке грунтується на порівнянні структури укладки вірусних білків, проте пропонує кілька нових доказів. Суперсімейки укладки білків Fold Super Families (FSF) — це білки, які мають схожі структури укладки поліпептидного ланцюга, незалежно від фактичної послідовності їх амінокислот, і вони, як було виявлено, можуть слугувати доказом філогенії вірусів. Протеом вірусу, вірусний протеом, все ще містить сліди давньої історії еволюції, які можна вивчити сьогодні. Дослідження суперсімейки укладки бліків свідчить про існування давніх клітинних ліній, спільних як для клітин, так і для вірусів, до появи «останнього універсального клітинного предка», що породив сучасні клітини. Еволюційний тиск на зменшення геному та розміру частинок, можливо, з часом призвів до зменшення клітин вірусів до сучасних вірусів, тоді як інші співіснуючі клітинні лінії з часом еволюціонували до сучасних клітин.[13] Крім того, велика генетична відстань між РНК та ДНК FSF свідчить про те, що гіпотеза РНК світу може мати нові експериментальні докази з тривалим проміжним періодом еволюції клітинного життя.
Остаточне виключення гіпотези про походження вірусів важко зробити на Землі, оскільки віруси та клітини сьогодні повсюдно взаємодіють між собою, і древні гірські породи, в яких могли бути знайдені сліди вірусів, на Землі, зустрічаються дуже рідко або вже відсутні. Тому з астробіологічної точки зору вже пропонувалось, щоб на небесних тілах, таких як Марс, слід активно шукати не тільки клітини, а й сліди колишніх віріонів або вірусів. Якщо на іншому небесному тілі знайдуть сліди віріонів, але жодних клітин, це буде вагомим свідченням першої гіпотези про вірус.[14]
Еволюція
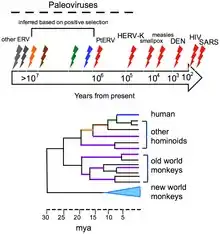
Віруси не утворюють скам'янілостей у традиційному розумінні, оскільки вони набагато менші, ніж найдрібніші колоїдні частинки, що утворюють осадові гірські породи, які призводять до фосилізації рослин та тварин. Однак геноми багатьох організмів містять ендогенні вірусні елементи (ЕВЕ). Ці ДНК послідовності являються залишками древніх вірусних генів і геномів, які «вторглись» в клітини зародкової лінії хазяїна. Наприклад, геноми більшості видів хребетних містять сотні-тисячі послідовностей, отриманих від древніх ретровірусів. Ці ендогенні вірусні елементи являються цінним джерелом ретроспективних даних по еволюційній історії вірусів і породили науку палеовірусологію. Еволюційну історію вірусів можна в деякій мірі вивести із аналізу сучасних вірусних геномів. Були виміряні швидкості мутацій для багатьох вірусів, а використання молекулярного годинника дозволяє визначити терміни розходження.[15]
Еволюційну історію вірусів можна певною мірою зробити висновком з аналізу сучасних вірусних геномів. Вимірювали швидкість мутації багатьох вірусів, і застосування молекулярного годинника дозволяє визначити терміни розбіжностей.[16]
Віруси розвиваються шляхом змін старих або набуттям нових послідовностей в їх РНК (або ДНК), деякі досить швидко, і найкраще адаптовані мутанти швидко перевершують чисельно своїх менш підходящих аналогів. У цьому сенсі їх еволюція є Дарвінівською.[18] Спосіб розмноження вірусів у клітинах-хазяїнах робить їх особливо сприйнятливими до генетичних змін, які пришвидшують їхню еволюцію.[18] РНК-віруси особливо схильні до мутацій.[19] У клітинах-хазяїнах існують механізми виправлення помилок під час реплікації ДНК, і вони починають діяти, коли клітини діляться.[19] Ці важливі механізми запобігають передачі нащадкам потенційно летальних мутацій. Але ці механізми не працюють для РНК, і коли РНК-вірус реплікується у своїй клітині-хазяїні, зміни в їх генах час від часу вносяться помилково, деякі з них є летальними. Одна частинка вірусу може продукувати мільйони вірусів-нащадків лише за один цикл реплікації, тому поява кількох «дефектних» вірусів не є проблемою. Більшість мутацій є «тихими» і не призводять до явних змін у потомстві вірусів, однак деякі надають переваги, які підвищують їх пристосованість до умов навколишнього середовища. Це можуть бути зміни вірусних часточок, які маскують їх від ідентифікації клітинами імунної системи, або зміни, що роблять противірусні препарати менш ефективними. Обидві ці зміни часто трапляються з ВІЛ.[20]
Багато вірусів (наприклад, вірус грипу А) можуть «перетасовувати» свої гени з іншими вірусами, коли два однакові штами заражають одну і ту ж клітину. Це явище називається антигенним зсувом і часто є причиною появи нових і більш вірулентних штамів. Інші віруси змінюються повільніше, оскільки мутації в їх генах поступово накопичуються з часом — процес, відомий як антигенний дрейф.[21]
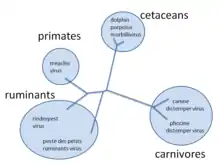
Завдяки цим механізмам нові віруси постійно з'являються і являють собою постійний виклик спробам контролювати хвороби, які вони викликають.[22][23] Зараз відомо, що більшість видів вірусів мають спільних предків, і хоча гіпотеза «первинності вірусів» ще не отримала повного визнання, немає ніяких сумнівів в тому, що тисячі видів сучасних вірусів еволюціонували з менш численних древніх.[24] Наприклад, морбілівіруси — це група тісно пов'язаних, але різних вірусів, які заражають широкий спектр тварин. До групи входить вірус кору, який заражає людей і приматів; вірус собачої чуми, який заражає багатьох тварин, включаючи собак, котів, ведмедів, ласок і гієн; чума великої рогатої худоби, яка заразила велику рогату худобу та буйволів; та інші віруси тюленів, морських свиней та дельфінів.[17] Хоча неможливо довести, який із цих вірусів, що швидко розвиваються є найбільш раннім, оскільки така тісно пов'язана група вірусів, що зустрічається у таких різноманітних господарів, передбачає можливість існування спільного предка в минулому.[17]
Бактеріофаг
Вірус ешерихії Т4 (фаг Т4) — це вид бактеріофагів, що інфікує Кишкову паличку. Це дволанцюжковий вірус ДНК сімейства Myoviridae. Фаг Т4 — являється облігатним внутрішньоклітинним паразитом, який розмножується всередині бактеріальної клітини хазяїна, і його потомство вивільняється, коли хазяїн руйнується лізисом. Повна послідовність геному фага Т4 кодує близько 300 генних продуктів.[25] Ці вірулентні віруси є одними з найбільших, найскладніших вірусів, які відомі, та одними з найбільш досліджених модельних організмів. Вони зіграли ключову роль у розвитку вірусології та молекулярної біології. Число зареєстрованих генетичних гомологій між фагами Т4 та бактеріями, а також між фагами Т4 та еукаріотами є подібними, що свідчить про те, що фаг Т4 має спільне походження з бактеріями та еукаріотами і має приблизно однакову схожість з кожним із них.[26] Фаг Т4 міг відрізнятися в процесі еволюції від загального предка бактерій та еукаріотів або від рано еволюціонуючого члена будь-якої лінії. Більшість фагових генів, що демонструють гомологію з бактеріями та еукаріотами, кодують ферменти, що діють у всюдисущих процесах реплікації ДНК, репарації ДНК, рекомбінації та синтезу нуклеотидів.[26] Ці процеси, ймовірно, розвивалися дуже рано. Можливо, адаптаційні особливості ферментів, що каталізують ці ранні процеси, підтримувались у фагах Т4, бактеріальних та еукаріотичних ліній, оскільки вони вже були добре перевіреними для вирішення основних функціональних проблем на той час, коли ці лінії розходилися.
Передача
Завдяки еволюції віруси змогли продовжувати своє інфекційне існування. Швидкі темпи мутацій та природний відбір дали вірусам переваги подальшого поширення. Одним із способів поширення вірусів є еволюція механізму передачі вірусів. Вірус може знайти нового господаря через:[27]
- Крапельну передачу — передається через біологічні рідини (чхання на когось)
- Прикладом є вірус грипу.[28]
- Передача повітряно-крапельним шляхом- передається по повітрю (передається дихальними шляхами).
- Прикладом може бути передача вірусного менінгіту.[29]
- Векторна передача — підхоплюється носієм і передається новому хазяїну.
- Прикладом може служити вірусний енцефаліт[30]
- Передача через воду — залишаючи господаря, заражаючи воду та споживанням разом з водою новим господарем.
- Поліовірус являється прикладом цього.[31]
- Повільна передача — вірус живе довгий час поза організмом хазяїна.
- Вірус віспи також є прикладом цього[31]
Існує також кілька ідей, пов'язаних з ідеєю, що вірулентність або шкода, яку завдає вірус своєму господареві, залежить від кількох факторів. Ці фактори також впливають на те, як з часом змінюватиметься рівень вірулентності. Віруси, які передаються шляхом вертикальної передачі (передача потомству господаря), в процесі еволюції будуть мати нижчий рівень вірулентності. Віруси, які передаються шляхом горизонтальної передачі (передача між представниками одного виду, які не знаходяться в родинних стосунках), як правило, будуть мати більш високий ступінь вірулентності.[32]
Див. також
- Катастрофа помилок
- Квазівиди вірусів
- Класифікація вірусів
- ДНК-віруси
- РНК-віруси
- Філодинаміка вірусів
- Хронологія людини
- Хронологія природи
Примітки
- Mahy, B. W. J.; Van Regenmortel, M. H. V. (2010). Desk encyclopedia of general virology. Oxford, UK: Academic Press. с. 24. ISBN 0-12-375162-4. OCLC 480587729.
- Villarreal, Luis P. (2005). Viruses and the evolution of life. Washington, D.C.: ASM Press. ISBN 1-55581-309-7. OCLC 57575788.
- Mahy, B. W. J.; Van Regenmortel, M. H. V. (2010). Desk encyclopedia of general virology. Oxford, UK: Academic Press. с. 25. ISBN 0-12-375162-4. OCLC 480587729.
- Mahy, B. W. J.; Van Regenmortel, M. H. V. (2010). Desk encyclopedia of general virology. Oxford, UK: Academic Press. с. 26. ISBN 0-12-375162-4. OCLC 480587729.
- Dimmock, N. J.; Leppard, K. N. (2007). Introduction to modern virology (вид. 6th ed). Malden, MA: Blackwell Pub. с. 16. ISBN 1-4051-3645-6. OCLC 65207057.
- Krupovic, Mart; Dolja, Valerian V.; Koonin, Eugene V. (2019-07). Origin of viruses: primordial replicators recruiting capsids from hosts. Nature Reviews Microbiology (англ.) 17 (7). с. 449–458. ISSN 1740-1526. doi:10.1038/s41579-019-0205-6. Процитовано 1 березня 2021.
- Nasir, Arshan; Kim, Kyung Mo; Caetano-Anollés, Gustavo (2012-09). Viral evolution: Primordial cellular origins and late adaptation to parasitism. Mobile Genetic Elements (англ.) 2 (5). с. 247–252. ISSN 2159-256X. PMC PMC3575434. PMID 23550145. doi:10.4161/mge.22797. Процитовано 1 березня 2021.
- Topley, W. W. C.; Collier, L. H.; Balows, Albert; Sussman, Max; Topley, W. W. C. (1998). Topley & Wilson's microbiology and microbial infections. (вид. 9th ed.). London: Arnold. с. 11. ISBN 0-340-61470-6. OCLC 38290809.
- Topley, W. W. C.; Collier, L. H.; Balows, Albert; Sussman, Max; Topley, W. W. C. (1998). Topley & Wilson's microbiology and microbial infections. (вид. 9th ed.). London: Arnold. с. 11–12. ISBN 0-340-61470-6. OCLC 38290809.
- Mahy, B. W. J.; Van Regenmortel, M. H. V. (2010). Desk encyclopedia of general virology. Oxford, UK: Academic Press. с. 362–378. ISBN 0-12-375162-4. OCLC 480587729.
- Forterre, Patrick (2010). Giant Viruses: Conflicts in Revisiting the Virus Concept. Intervirology (англ.) 53 (5). с. 362–378. ISSN 1423-0100. doi:10.1159/000312921. Процитовано 1 березня 2021.
- Piast, Radosław W. (2019-06). Shannon's information, Bernal's biopoiesis and Bernoulli distribution as pillars for building a definition of life. Journal of Theoretical Biology (англ.) 470. с. 101–107. doi:10.1016/j.jtbi.2019.03.009. Процитовано 1 березня 2021.
- Nasir, Arshan; Caetano-Anollés, Gustavo (2015-09). A phylogenomic data-driven exploration of viral origins and evolution. Science Advances (англ.) 1 (8). с. e1500527. ISSN 2375-2548. PMC PMC4643759. PMID 26601271. doi:10.1126/sciadv.1500527. Процитовано 1 березня 2021.
- Janjic, Aleksandar (2018-12). The Need for Including Virus Detection Methods in Future Mars Missions. Astrobiology (англ.) 18 (12). с. 1611–1614. ISSN 1531-1074. doi:10.1089/ast.2018.1851. Процитовано 1 березня 2021.
- Emerman, Michael; Malik, Harmit S. (9 лютого 2010). Paleovirology—Modern Consequences of Ancient Viruses. У Virgin, Skip W. PLoS Biology (англ.) 8 (2). с. e1000301. ISSN 1545-7885. PMC PMC2817711. PMID 20161719. doi:10.1371/journal.pbio.1000301. Процитовано 1 березня 2021.
- Lam, Tommy Tsan-Yuk; Hon, Chung-Chau; Tang, Julian W. (2010-02). Use of phylogenetics in the molecular epidemiology and evolutionary studies of viral infections. Critical Reviews in Clinical Laboratory Sciences (англ.) 47 (1). с. 5–49. ISSN 1040-8363. doi:10.3109/10408361003633318. Процитовано 1 березня 2021.
- Barrett, Thomas; Pastoret, Paul-Pierre; Taylor, William P.; Scott, Gordon; Provost, A. (2006). Rinderpest and peste des petits ruminants : virus plagues of large and small ruminants. London: Academic. ISBN 978-0-12-088385-1. OCLC 162569523.
- Dimmock, N. J.; Leppard, K. N. (2007). Introduction to modern virology (вид. 6th ed). Malden, MA: Blackwell Pub. ISBN 1-4051-3645-6. OCLC 65207057.
- Domingo, Esteban; Escarmís, Cristina; Sevilla, Noemi; Moya, Andres; Elena, Santiago F.; Quer, Josep; Novella, Isabel S.; Holland, John J. (1996-06). Basic concepts in RNA virus evolution. The FASEB Journal (англ.) 10 (8). с. 859–864. ISSN 0892-6638. doi:10.1096/fasebj.10.8.8666162. Процитовано 1 березня 2021.
- Boutwell, Christian L.; Rolland, Morgane M.; Herbeck, Joshua T.; Mullins, James I.; Allen, Todd M. (15 жовтня 2010). Viral Evolution and Escape during Acute HIV‐1 Infection. The Journal of Infectious Diseases (англ.) 202 (S2). с. S309–S314. ISSN 0022-1899. PMC PMC2945609. PMID 20846038. doi:10.1086/655653. Процитовано 1 березня 2021.
- Chen, Jiezhong; Deng, Yi-Mo (2009). Influenza virus antigenic variation, host antibody production and new approach to control epidemics. Virology Journal (англ.) 6 (1). с. 30. ISSN 1743-422X. PMC PMC2666653. PMID 19284639. doi:10.1186/1743-422X-6-30. Процитовано 1 березня 2021.
- Fraile, Aurora; García-Arenal, Fernando (2010). The Coevolution of Plants and Viruses. Advances in Virus Research (англ.) 76. Elsevier. с. 1–32. ISBN 978-0-12-374525-5. doi:10.1016/s0065-3527(10)76001-2.
- Tang, Julian W.; Shetty, Nandini; Lam, Tommy T.Y.; Hon, K.L. Ellis (2010-09). Emerging, Novel, and Known Influenza Virus Infections in Humans. Infectious Disease Clinics of North America (англ.) 24 (3). с. 603–617. PMC PMC7127320. PMID 20674794. doi:10.1016/j.idc.2010.04.001. Процитовано 1 березня 2021.
- Mahy, B. W. J.; Van Regenmortel, M. H. V. (2010). Desk encyclopedia of general virology. Oxford, UK: Academic Press. ISBN 0-12-375162-4. OCLC 480587729.
- Miller, Eric S.; Kutter, Elizabeth; Mosig, Gisela; Arisaka, Fumio; Kunisawa, Takashi; Rüger, Wolfgang (2003-03). Bacteriophage T4 Genome. Microbiology and Molecular Biology Reviews (англ.) 67 (1). с. 86–156. ISSN 1092-2172. PMC PMC150520. PMID 12626685. doi:10.1128/MMBR.67.1.86-156.2003. Процитовано 1 березня 2021.
- Bernstein, H; Bernstein, C (1989). Bacteriophage T4 genetic homologies with bacteria and eucaryotes.. Journal of Bacteriology (англ.) 171 (5). с. 2265–2270. ISSN 0021-9193. doi:10.1128/JB.171.5.2265-2270.1989. Процитовано 1 березня 2021.
- "Evolution from a virus's view" (англ.). 27 листопада 2017. Процитовано 27 листопада 2017.
- "Key Facts About Influenza (Flu)". Процитовано 16 жовтня 2017.
- "Meningitis, Viral". Процитовано 4 грудня 2017.
- Encephalitis. Процитовано 5 грудня 2017.
- Smallpox. Процитовано 13 липня 2017.
- Implications of horizontal and vertical pathogen transmission for honey bee epidemiology.
Література
- (англ.) Barrett, Thomas C; Pastoret, Paul-Pierre; Taylor, William J. (2006). Rinderpest and peste des petits ruminants: virus plagues of large and small ruminants. Amsterdam: Elsevier Academic Press. ISBN 0-12-088385-6.
- (англ.) Leppard, Keith; Dimmock, Nigel; Easton, Andrew (2007). Introduction to Modern Virology. Blackwell Publishing Limited. ISBN 978-1-4051-3645-7.
- (англ.) Mahy, W.J.; Van Regenmortel, MHV, ред. (2009). Desk Encyclopedia of General Virology. Academic Press. ISBN 978-0-12-375146-1.
- (англ.) Sussman, Max; Topley, W.W.C.; Wilson, Graham K.; Collier, L.H.; Balows, Albert (1998). Topley & Wilson's microbiology and microbial infections. Arnold. ISBN 0-340-66316-2.
Посилання
- (англ.) Where Did Viruses Come From?. PBS Eons. 12 червня 2018.
