Філогенетичне дерево
Філогенети́чне де́рево (еволюційне дерево, дерево життя) — дерево, що відображає еволюційні взаємозв'язки між різними видами, іншими таксонами, генами або іншими об'єктами, що мають загального предка.
| Частина серії статей на тему: |
| Еволюційна біологія |
|---|
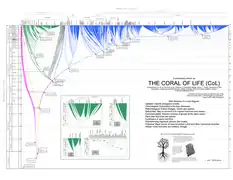 «Корал життя» — сучасне представлення філогенетичного древа всього живого на планеті |
|
|
Процеси |
|
Розвиток теорії
|
|
Дисципліни і застосування
|
|
Вплив на соціум
|
|
|
Вершини філогенетичного дерева діляться на три класи: листя, вузли і (максимум один) корінь. Листя — це кінцеві вершини, тобто ті, в які входять рівно по одному ребру; кожен лист відображає деякий вид живих організмів (або інший об'єкт, схильний до еволюції, наприклад білковий домен). Кожен вузол представляє еволюційну подію: розділення предкового виду на два або більше, які надалі еволюціонували незалежно. Корінь представляє загального предка всіх даних об'єктів. Ребра дерева філогенезу прийнято називати «гілками».
Ідея «дерева» з'явилася в ранніх поглядах на життя, як на процес розвитку від простих форм до складних. Сучасні еволюційні біологи продовжують використовувати дерева для ілюстрації еволюції, оскільки воно наочно показує розвиток і походження видів.
Типи філогенетичних дерев
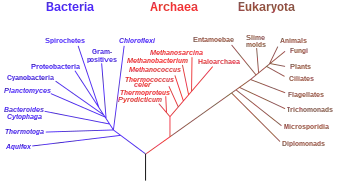
Вкорінене дерево — дерево, що містить виділену вершину — корінь. Вкорінене дерево можна вважати орієнтованим графом, оскільки воно має природну орієнтацію — від кореня до листя. Кожен вузол вкоріненого дерева відповідає останньому загальному предкові листя дерева, що лежить нижче. На малюнку представлене вкорінене філогенетичне дерево, забарвлене відповідно до деменам живих організмів у «системі трьох доменів»[2].
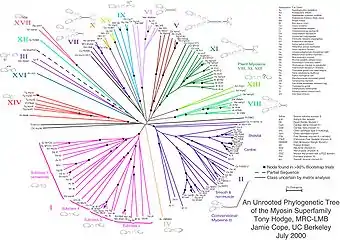
Невкорінене дерево не містить кореня і відображає зв'язок листя без передбачуваного положення загального предка. Необхідність розглядати невкорінені дерева виникає через те, що часто зв'язки між вузлами відновити легше, ніж напрямок еволюції. На другому малюнку показано невкорінене філогетичне дерево[3]. Найдостовірнішим методом для перетворення невкоріненого дерева на вкорінене (для цього треба або оголосити коренем один з вузлів, або розбити одну з гілок дві, що виходять з кореня) є використання достовірної «зовнішньої групи» видів — достатньо близьких до набору видів, що цікавить нас, але в той же час явний є окремою групою. В інших випадках положення кореня можна встановити, виходячи з додаткових припущень про швидкість еволюції досліджуваних об'єктів.
Як вкорінене, так і невкорінене філогенетичні дерева можуть бути біфуркаційними або мультифуркаційними, а також маркованими або немаркованими. У біфуркаційному дереві до кожного вузла підходять рівно три гілки (у разі вкоріненого дерева — одна вхідна гілка і дві вихідні). Таким чином біфуркаційне дерево припускає, що всі еволюційні події полягали в походженні від предкового об'єкта рівно двох нащадків. До вузла мультифуркаційного дерева можуть підходити чотири й більше гілок. Марковане дерево містить назви листків, тоді як немарковане просто відображає топологію.
Число можливих дерев для наданого ряду вершин-листів залежить від виду дерева, але завжди існує більше можливих мультифуркаційних дерев ніж біфуркаційних, більше маркірованих ніж немаркірованих і більше вкорінених, ніж невкорінених. Остання відмінність має найбільше біологічне значення — існує багато місць на невкоріненому дереві, куди можна помістити корінь. Для маркірованих біфуркаційних дерев існує

вкорінених дерев

невкорінених дерев, де n представляє число вузлів. Число невкорінених дерев для n вузлів дорівнює числу вкорінених для n-1 вузлів[4].
Дендрограма — загальний термін, що позначає схематичне представлення філогенетичного дерева.
Кладограма — філогенетичне дерево, що не містить інформації про довжини гілок.
Філограма (або фенограма) — філогенетичне дерево, що містить інформацію про довжини гілок — ці довжини представляють зміну якоїсь характеристики.
Хронограмма — філограма, довжини гілок в якій представляють еволюційний час.
Будівництво філогенетичних дерев
Філогенетичні дерева, що складаються з нетривіального числа вхідних послідовностей побудовані з використанням обчислювальних Філогенетичних методів. Дистанційні методи матрицею, таких як UPGMA, які розраховують генетичну відстань від множинного вирівнювання послідовностей, але не посилаються на еволюційну модель. Багато методів вирівнювання послідовності, такі як ClustalW також створюють дерева за допомогою простих алгоритмів (тобто ті, які засновані на відстані) в побудові дерева.
Максимальна скупість ще один простий спосіб оцінки філогенетичних дерев, але припускає неявну модель еволюції (тобто скупість). Більш сучасні методи використовують критерій оптимальності в максимальній ймовірності.
Більш сучасні методи використовують критерії оптимальності в максимальній ймовірності, часто в Байєсівській структурі, і застосовують явну модель еволюції до оцінки філогенетичного дерева.
Методи побудови філогенетичних дерев можуть бути оцінені на основі декількох критеріїв: [ 5]
- ефективність (скільки пам'яті потрібно, щоб обчислити відповідь?)
- потужність (чи дані використовуються ефективно чи даремно?)
- консистенція (чи буде сходитися на кожен раз відповідь, якщо кожен раз даються різні дані по тій же модельній задачі?)
- надійність (чи добре справляються з порушеннями припущень базової моделі?)
- фальсифікованість (якщо теорія не може бути фальсифікована, немає сенсу навіть дослідження доказів).
Методи побудови філогенетичних дерев також привернули увагу математиків. Дерева також можуть бути побудовані з використанням T-теорії. [ 6]
Див. також
Примітки
- Hodge, T. & M. J. T. V. Cope. 2000. A Myosin Family Tree. Journal of Cell Science 113: 3353-3354. See also the Myosin external link below.
- Woese, C. R.. 1998. The Universal Ancestor. Proceedings of the National Academy of Sciences 95: 6854-6859.
- Maher, B. A. 2002. Uprooting the Tree of Life. The Scientist 16: 18 (Sep. 16, 2002); subscription only
- Felsenstein J. (2004). Inferring Phylogenies Sinauer Associates: Sunderland, MA.
Посилання
- РОДОВІДНЕ ДЕРЕВО
- 5.Penny, D.; Hendy, M. D.; Steel, M. A. (1992). «Progress with methods for constructing evolutionary trees». Trends in Ecology and Evolution 7: 73–79. doi:10.1016/0169-5347(92)90244-6
- 6.A. Dress, K. T. Huber, and V. Moulton. 2001. Metric Spaces in Pure and Applied Mathematics. Documenta Mathematica LSU 2001: 121—139
