Кладистика
Кладистика (від дав.-гр. κλάδος (kládos) — гілка) — спеціальний підхід до біологічної класифікації, в рамках якого організми класифікують ґрунтуючись на порядку, у якому вони відгалужувались від еволюційного дерева, незважаючи на їхню морфологічну подібність[1]. Засновником та автором найбільшого внеску в дисципліну є німецький ентомолог Віллі Генніг, котрий називав її «філогенетичною систематикою» (Hennig, 1979), хоча зараз останній термін має ширше значення. Зараз кладистика все більше розглядається як основний підхід до біологічної класифікації, і всі сучасні класифікаційні системи тією або іншою мірою включають інформацію, отриману за допомогою цього підходу.
| Частина серії статей на тему: |
| Еволюційна біологія |
|---|
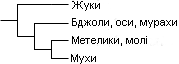 Приклад кладограми, що показує взаємовідносини між різними групами комах |
|
|
Процеси |
|
Розвиток теорії
|
|
Дисципліни і застосування
|
|
Вплив на соціум
|
|
|
Результатом кладистичного аналізу походження таксона є спеціальні діаграми, що називаються «кладограмами», які віддзеркалюють гіпотетичні філогенетичні зв'язки. Кладистичний аналіз може ґрунтуватись на матеріалі різного обсягу, доступному для конкретного дослідника. Наприклад, стосовно викопних організмів, як правило, доступні тільки морфологічні дані, тоді як для сучасних живих істот використовується також аналіз послідовностей ДНК (так звані «молекулярні дані») та біохімічні дані.
У кладограмі всі таксони розташовані на кінцях гілок, і кожне розгалуження є дихотомічним (двійковим). Два таксони, розділені одним розгалуженням, називаються сестринськими таксонами або сестринськими групами. Кожна частина загального дерева, незалежно від того, налічує вона один або тисячі таксонів, називається «клада». Клада включає всі таксони, які мають спільного унікального предка (такого, що не є попередником ніякого більше організму в діаграмі). Кожна клада характеризується набором ознак, притаманних її членам, але відсутніх у предків. Ці ознаки клади називаються «синапоморфії» (спільні риси). Наприклад, затверділа передня пара крил (надкрила) є синапоморфіями жуків, а кільцеве листоутворення (або розвертання вайїв) є синапоморфією папоротей.
Визначення
Характеристична ознака, наявна як у кореневого виду, так і у його нащадків, називається «плезіоморфія» («близька форма», також називається «попередниковою формою»). Характеристична ознака, що наявна тільки у пізніших нащадків, називається «апоморфія» (означає «відокремлена форма», також називається «похідною формою») для цієї групи. Прикметники «плезіоморфний» та «апоморфний» використовуються замість «примітивний» та «передовий» (або ж «просунутий») для уникнення оціночного судження про еволюційні форми, позаяк обидва вони можуть позначати структури, що є вкрай корисними за деяких обставин та шкідливими — за інших. Нерідко набір плезіоморфних форм неформально називається «базисом» для клади або клад, котрим ций набір притаманний.
Кілька інших поширених термінів кладистики введено для опису кладограм та позицій таксонів всередині них. Вид або клада є базальною відносно іншої клади, якщо перша має більше плезіоморфних рис, ніж друга. Звичайно базальна група налічує малу кількість видів порівняно з розвинутішими групами; наявність в кладограмі базальної групи не є обов'язковою. Наприклад, при спільній кладистичній класифікації птахів та ссавців жодна з цих груп не є базальною для іншої.
Клада або вид, що знаходиться в кладограмі всередині іншої клади, може називатись «вкладеною» або «вкладеним» в цю кладу.
Історія назви
Класифікаційний підхід, зараз відомий як кладистика, був надихнутий працями Віллі (Вільгельма) Генніга. Але його головна книга, навіть у редакції 1979 року, не містить власне терміну «кладистика». В цій книзі автор називає свій підхід філогенетичною систематикою, і цю ж назву отримала сама книга (Hennig, 1979). Оглядова стаття Дюпуа (Dupuis,1984) стверджує, що термін «клада» був запропонований в 1958 році Джуліаном Гакслі, «кладистика» — Кейном та Харрісом в 1960-му, а «кладист» (для позначення послідовника школи Генніга) — Майром в 1965.[2] Декотрі дискусії, куди були залучені кладисти, тривали з 19-го сторіччя, але кладисти приєднались до них пізніше і з новими поглядами, що описане у передмові до книги Генніга редакції 1979-го року, де Розен, Нельсон та Патерсон зазначали наступне[3]:
| Обтяжений неясними та ковзкими ідеями про адаптацію, еволюційну придатність, біологічні види та природний добір, неодарвінізм (що визначається як «еволюційний» підхід до біологічної систематики Майром та Сімпсоном) не має визначеного методу дослідження, а також прийшов до залежності від консенсусу авторитетів у питаннях еволюційної інтерпретації та класифікації. |
Методи кладистики
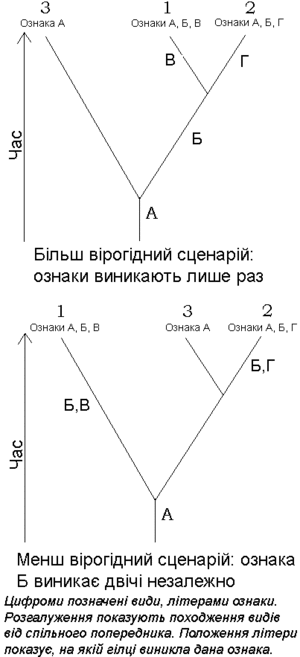
Масив інформації, що може бути підданий кладистичному аналізу, має бути організований спеціальним чином. Для цього, перш за все, треба провести розмежування між ознаками (або характеристиками) та їх станами (характеристичними станами). Наприклад, колір пір'їн може бути блакитним у одного виду та червоним у іншого. Тоді «блакитні пера» та «червоні пера» будуть двома станами однієї ознаки — «кольору пір'я».
Дослідник має визначити, яка ознака (або ознаки) була наявна до появи останнього спільного попередника (плезіоморфія), а яка була наявна у останнього спільного попередника (синапоморфія) шляхом виділення одного чи більше кореневих видів. Кореневий вид — це організм, котрий не належить до досліджуваної групи, але є близько з нею спорідненим. Це робить вибір кореневого виду важливою задачею, позаяк такий вибір здатний серйозно вплинути на структуру кладистичного дерева. Треба зауважити, що при характеристиці клад використовуються лише синапоморфії.[4]
Наступною стадією є укладання різних можливих кладограм та їхня перевірка. В ідеалі клади мають багато «узгоджених» синапоморфій; в такому ідеальному випадку очікують наявність достатньо великої кількості справжніх синаптоморф, котрі не можуть бути прихованими гомоплазіями, появу котрих спричинює конвергентна еволюція (тобто якостями, котрі відтворюють одна одну завдяки впливу навколишнього середовища або спільного функціонального використання, а не завдяки спільному походженню). Відомим прикладом гомоплазії, що виникла завдяки конвергентній еволюції, є ознака «наявність крил». Хоча крила птахів, кажанів та комах виконують однакові функції, кожне з цих крил еволюціонувало незалежно, що може бути простежено завдяки їхній анатомії. Якщо ж птах, кажан та крилата комаха будуть об'єднані ознакою «наявність крил», в масив даних буде штучно внесена гомоплазія, що зруйнує аналіз і скоріше за все призведе, в результаті, до невірної побудови еволюційної картини.
Застосування гомоплазії в морфологічних наборах даних часто можна уникнути шляхом якнайточнішого визначення характеристичних ознак та збільшення їхньої кількості: в попередньому прикладі, використовуючи ознаки «крила з пір'ям», «крила з хітиновим екзоскелетом» та «шкірясті крила» як характеристичні ознаки, можна уникнути еволюційно хибного об'єднання трьох перелічених груп тварин на основі гомоплазії. При аналізі «супердерев» (баз даних, що включають якнайбільше таксонів досліджуваної клади), застосування неточних ознак може стати неминучим, позаяк у зворотному випадку ознаки можуть стати незастосуваними для всіх чисельних таксонів. Скажімо, така ознака, як наявність крил, не може бути застосована для аналізу філогенії справжніх багатоклітинних тварин (Eumetazoa), так як більшості видів цього таксона дана ознака не притаманна. Таким чином, обережний вибір та визначення характеристичних ознак є іншим важливим елементом кладистичного аналізу. При помилковому визначенні кореневого виду та набору ознак ніякі методи побудови кладограм не зможуть дати в результаті філогенетичну систему, що відповідає еволюційній реальності.
Для практично будь-якого таксона можливо побудувати не одну, а велику кількість різних кладограм, базуючись на різних кореневих видах та наборах характеристичних ознак; але з них вибирають одну-єдину, керуючись принципом парсимонії: найкомпактнішу систему, котра, у сполученні з найменшими можливими змінами характеристичних ознак (синапоморфій), дає непротирічну картину походження клади (загалом, це є варіантом міркування згідно з бритвою Окама). Хоча на початку такий аналіз проводився «вручну», згодом для нього стали застосовуватись комп'ютери із спеціальним програмним забезпеченням, яке дозволяє оперувати на порядки більшими наборами даних та кількістю ознак. Такі програми (на кшталт PAUP та інших подібних) дозволяють робити статистичну оцінку ймовірності нод (вузлів, розгалужень) побудованої кладограми.
Важливо також зауважити, що ноди кладограми не обов'язково віддзеркалюють розходження еволюційних гілок, а натомість лише розходження станів ознак, що спостерігаються між цими гілками. Ознаки, що полягають у різниці послідовностей ДНК, здатні розходитись після того, як генетичний дрейф між популяціями редукується до деякої порогової величини, в той час як помітні морфологічні зміни, зазвичай будучі епістатичними (тобто результатом взаємодії кількох генів) виявляються лише після того, як таксони, що розійшлись, відокремлено еволюційно розвивались протягом деякого (зазвичай досить значного) часу; так, біологічні підвиди найчастіше можуть бути розрізнені генетично, але не морфологічно (за будовою тіла чи внутрішньою анатомією).
З огляду на те, що секвенування ДНК стає все дешевшим та легшим у впровадженні, молекулярна систематика набуває все більшої популярності як спосіб реконструкції філогенезу. Використання критерію парсимонії є лише одним з методів з'ясування філогенії таксона на базі молекулярно-біологічних даних; методи максимальної правдоподібності та Байєзівського виведення, котрі включають в себе докладно розроблені моделі еволюції послідовностей, є відомими не-геннігіанськими методами оцінки даних секвенування. Іншим потужним методом реконструкції філогенії є використання ретротранспозонних маркерів геному, котрі вважають такими, що менш піддаються впливу реверсії секвенованих даних, котра (реверсія) може зробити дані непридатними для побудови філогенетичної послідовності. Також перевагою ретротранспозонних маркерів є те, що вони мало піддаються гомоплазії через інтегрування в геном у повністю випадкових місцях (хоча така статистична випадковість, щонайменше подекуди, є сумнівною).[5]
В ідеалі морфологічні, молекулярно-біологічні, та інші (поведінкові, екологічні, палеонтологічні тощо) філогенетичні дані мають бути узагальненими при опрацюванні підсумкового висновку; при цьому ніякий з методів не є доказовішим за інші, але всі вони мають різні внутрішні джерела помилок. Наприклад, конвергенція розвитку ознак (гомоплазія) набагато частіше з'являється при аналізі морфологічних даних, ніж серед даних, отриманих при молекулярному секвенуванні, але реверсії ознак в них обох зустрічаються з приблизно однаковою частотою; зазвичай, морфологічні гомоплазії можуть бути викриті при достатньо уважному та детальному аналізі характеристичних ознак.
Кладистика не прив'язана до жодної з наявних теорій еволюції, беручи до уваги лише загальну інформацію про походження з (часом) істотними її модифікаціями. Завдяки цьому кладистичні методи можуть бути (і бувають) використаними в небіологічних дисциплінах, включаючи історичну лінгвістику та завдання з визначення авторства текстів.
Кладистична класифікація
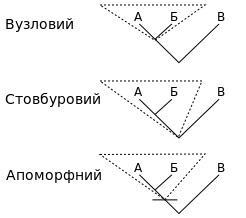
Вузловий: найнедавніший спільний попередник видів А та Б і всі його нащадки
Стовбурний: всі нащадки найдавнішого спільного попередника А та Б, що одночасно не є попередниками В
Апоморфний: Найдавніший спільний попередник А та Б, у якого наявна деяка визначальна апоморфія, та всі його нащадки
З 1960-х у біології виникла тенденція включати в класифікацію лише клади, відома під назвою «кладизм» або «кладистична таксономія». Іншими словами, кладисти вважають, що біологічна класифікація має бути реформована таким чином, щоб усунути всі не-клади. На відміну від кладистів, всі інші таксономісти наполягають на тому, що кожна з таксономічних груп живих істот віддзеркалює філогенетичну історію. Останні часто використовують кладистичний підхід, але також дозволяють в формуванні класифікаційних дерев використання як монофілетичних (що збігається з кладистичним підходом), так і парафілетичних таксонів. Як результат, вже як мінімум з початку XX століття роди та таксони нижчого рівня формували, базуючись на монофілетичному підході, в той час як таксони вищого рангу можуть бути (а такі як клас і вище — зазвичай і є) парафілетичними.
У цій системі впорядкування монофілетичною групою вважається клада, що складається з попередника та всіх його нащадків, які формують одну й лише одну еволюційну групу. Парафілетичною є така сама, але без деяких нащадків, які зазнали істотних змін. Наприклад, традиційний клас Плазуни не включає птахів, хоча птахи походять від плазунів; аналогічно, традиційна група Безхребетні є парафілетичною, тому що не включає хребетних, що походять від неї.
Група, що складається з членів, які походять з різних еволюційних ліній, називається поліфілетичною. Наприклад, сформована колись таксономічна група «Товстошкірі» (Pachydermata) була потім визнана поліфілетичною, позаяк включені до неї слони, носороги та бегемоти походять від різних, не споріднених між собою попередників. Аналогічно сформований колись тип Кишковопорожнинні був пізніше розділений на Кнідарій та Гребневиків на підставі їхнього походження від різних попередників та виниклих внаслідок цього істотних морфологічних розбіжностей. Еволюційні таксономісти вважають поліфілетичні групи помилками класифікації, спричиненими конвергенцією та іншими видами гомоплазії, що помилково інтерпретуються як гомології.
Наслідуючи Генніга, кладисти вважають, що парафілія є не менш неприйнятною для класифікації, аніж поліфілія. В рамках кладистики вважається, що монофілетичні групи можуть бути об'єктивно та строго визначеними, базуючись на спільних попередниках або синаптоморфах. На відміну від них, парафілетичні та поліфілетичні групи виділяються на основі аналізу ключових характеристик, у ході якого ступінь важливості цих характеристик є досить суб'єктивною. Кладисти зауважують, що, таким чином, це призводить до «градистичного» мислення, яке в свою чергу тяжіє до телеології, котру зазвичай вважають неприйнятною у еволюційних дослідженнях, оскільки телеологія має на увазі існування загального плану еволюції, котрий не може бути показаним експериментально.
Продовжуючи цю лінію критики, деякі кладисти стверджують, що таксони вищого за види рангу є забагато суб'єктивними, і тому їх виділення не репрезентує ніякої змістовної інформації. На основі цього, як стверджують такі критики, ці таксони не повинні використовуватись в систематиці взагалі. Таким чином, біологічна систематика, в їхньому розумінні, має повністю відійти від ліннеївської таксономії и стати простою ієрархією клад. Вагомість цієї системи аргументів критично залежить від того, як часто в ході еволюції переривається градуалістська рівновага екологічних умов. Квазі-стабільний стан екосистем призводить до повільного філогенезу, котрий може бути важко інтерпретованим у рамках ліннеївської систематики; в той же час катастрофічна подія виводить таксон з екологічної рівноваги і зазвичай призводить до розділення ознак клад. Це розділення відбувається протягом еволюційно короткого часу, і в результаті дає систему таксонів, що легко описується в рамках ліннеївської класифікації.[6]
Інші еволюційні систематики стверджують, що всі таксони за своєю сутністю є суб'єктивними, навіть коли вони віддзеркалюють еволюційні взаємовідносини, позаяк живі організми формують безперервне еволюційне дерево. В такому випадку, будь-яка лінія розділу є штучною і створює монофілетичний виділ над парафілетичним. Парафілетичні таксони є необхідними для класифікації більш ранніх секцій дерева — наприклад, ранні хребетні, що через деякий час розвинулись в родину Гомініди, не можуть бути вміщені більше в жодну монофілетичну родину. Також у цій системі аргументів наводиться те, що парафілетичні таксони надають інформацію про суттєві зміни в морфології, екології та еволюційній історії організмів — коротко кажучи, що і таксони, і клади є цінними для побудови правдивої картини систематики живої природи, але і ті, і ті — з деякими обмеженнями.
У полеміці навколо придатності кладистичних методів при побудові біологічної класифікації термін «монофілія» часто використовується у своєму старому значенні, де він включає в себе парафілію, поряд з чим використовується термін «голофілія» для опису «чистих» клад («монофілія» в геннігівському розумінні). Як приклад ненауковості емпіричного методу використовується наступний: якщо якась парафілетична еволюційна лінія приводить до виникнення клади через значну адаптивну радіацію і численні, добре помітні та важливі синапоморфії, особливо такі, що раніше в цій групі живих істот не зустрічалися, парафілія не визнається достатнім аргументом проти виділення таксона в ліннеївській системі класифікації (але це вважається достатнім контраргументом в рамках філогенетичної номенклатури). Наприклад, як було вказано вище, і зауропсиди («рептілії»), і птахи в ліннеївській систематиці мають однаковий ранг класів, хоча останні еволюційно є вкрай спеціалізованою гілкою рептилій.
Формальний збірник філогенетичної номенклатури (ФілоКод) зараз[коли?] знаходиться в розробці для пристосування його до кладистичної таксономії. Його планують використовувати і ті, хто намагається повністю уникати ліннеївської номенклатури, і ті, хто використовує в систематиці таксони разом із кладами. За деяких обставин (наприклад, при класифікації гесперорнісів) методи ФілоКоду було залучено для уникнення непевностей, що виникали при застосуванні ліннеївської систематики до цього таксона; таким чином, комбінація цих двох систем дає змогу сформувати таксономічну картину, котра розміщує групу живих істот на еволюційному дереві у повній відповідності до наявної наукової інформації.
Примітки
- Salvador Edward Luria, Stephen Jay Gould, Sam Singer. A view of life. Benjamin/Cummings Pub. Co., 1981 (англ.)
- Dupuis, C (1984). Willi Henning's Impact on Taxonomic Thought. Annual Review of Ecology and Systematics 15 (1): 1–25. ISSN 0066-4162. doi:10.1146/annurev.es.15.110184.000245.(англ.)
- Phylogenetic systematics / Willi Hennig; translated by D. Dwight Davis and Rainer Zangerl; foreword by Donn E. Rosen, Gareth Nelson, and Colin Patterson. Urbana: University of Illinois Press, 263 pages (1979)(англ.)
- Ковблюк, 2008, с. 95-96.
- Shedlock, Andrew M.; Okada, Norihiro (2000). SINE insertions: powerful tools for molecular systematics. BioEssays 22 (2): 148–160. ISSN 02659247. doi:10.1002/(SICI)1521-1878(200002)22:2<148::AID-BIES6>3.0.CO;2-Z.(англ.)
- de Queiroz, Kevin; Gauthier, Jacques (1992). Phylogenetic Taxonomy. Annual Review of Ecology and Systematics 23 (1): 449–480. ISSN 0066-4162. doi:10.1146/annurev.es.23.110192.002313.
Джерела
- Ковблюк М. М. (2008). Основи зоологічної номенклатури та систематики: Навчальний посібник для студентів біологічних спеціальностей вищих навчальних закладів. Сімферополь: ДІАЙПІ. с. 94-95. с. 148. ISBN 978-966-491-028-3.
- Ashlock, P D (1974). The Uses of Cladistics. Annual Review of Ecology and Systematics 5 (1): 81–99. ISSN 0066-4162. doi:10.1146/annurev.es.05.110174.000501.(англ.)
- Cuénot, Lucien (1940). «Remarques sur un essai d'arbre généalogique du règne animal». Comptes Rendus de l'Académie des Sciences de Paris 210: 23-27. Available free online at http://gallica.bnf.fr (No direct URL). This is the paper credited by *Hennig (1979) for the first use of the term 'clade'.
- de Queiroz, Kevin; Gauthier, Jacques (1992). Phylogenetic Taxonomy. Annual Review of Ecology and Systematics 23 (1): 449–480. ISSN 0066-4162. doi:10.1146/annurev.es.23.110192.002313.(англ.)
- Dupuis, C (1984). Willi Henning's Impact on Taxonomic Thought. Annual Review of Ecology and Systematics 15 (1): 1–25. ISSN 0066-4162. doi:10.1146/annurev.es.15.110184.000245.(англ.)
- Felsenstein, Joseph (2004). Inferring phylogenies. Sunderland, MA: Sinauer Associates. ISBN 0-87893-177-5.
- Hamdi, Hamdi; Hitomi Nishio, Rita Zielinski and Achilles Dugaiczyk (1999). «Origin and phylogenetic distribution of Alu DNA repeats: irreversible events in the evolution of primates». Journal of Molecular Biology 289: 861—871. PMID 10369767.
- Hennig, Willi (1950). Grundzüge einer Theorie der Phylogenetischen Systematik. Berlin: Deutscher Zentralverlag. .
- Hennig, Willi (1982). Phylogenetische Systematik (ed. Wolfgang Hennig). Berlin: Blackwell Wissenschaft. ISBN 3-8263-2841-8.
- Hennig, Willi (1975). «'Cladistic analysis or cladistic classification': a reply to Ernst Mayr». Systematic Zoology 24: 244—256. The paper he was responding to is reprinted in Mayr (1976).
- Hennig, Willi (1966). Phylogenetic systematics (tr. D. Dwight Davis and Rainer Zangerl). Urbana, IL: Univ. of Illinois Press (reprinted 1979 and 1999). ISBN 0-252-06814-9. Translated from manuscript and so never published in German.
- Hull, David L. (1979). «The limits of cladism». Systematic Zoology 28: 416—440.
- Kitching, Ian J.; Peter L. Forey, Christopher J. Humphries and David M. Williams (1998). Cladistics: Theory and practice of parsimony analysis, 2nd ed., Oxford University Press. ISBN 0-19-850138-2.
- Luria, Salvador; Stephen Jay Gould and Sam Singer (1981). A view of life. Menlo Park, CA: Benjamin/Cummings. ISBN 0-8053-6648-2.
- Mayr, Ernst (1982). The growth of biological thought: diversity, evolution and inheritance. Cambridge, MA: Harvard Univ. Press. ISBN 0-674-36446-5.
- Mayr, Ernst (1976). Evolution and the diversity of life (Selected essays). Cambridge, MA: Harvard Univ. Press. ISBN 0-674-27105-X. Reissued 1997 in paperback. Includes a reprint of Mayr's 1974 anti-cladistics paper at pp. 433–476, «Cladistic analysis or cladistic classification.» This is the paper to which Hennig (1975) is a response.
- Patterson, Colin (1982). «Morphological characters and homology». Joysey, Kenneth A; A. E. Friday (editors) Problems in *Phylogenetic Reconstruction, London: Academic Press.
- Rosen, Donn; Gareth Nelson and Colin Patterson (1979), Foreword provided for Hennig (1979)
- Shedlock, Andrew M; Norihiro Okada (2000). «SINE insertions: Powerful tools for molecular systematics». Bioessays 22: 148—160. ISSN 0039-7989. PMID 10655034.
- Sokal, Robert R. (1975). «Mayr on cladism — and his critics». Systematic Zoology 24: 257—262.
- Swofford, David L.; G. J. Olsen, P. J. Waddell and David M. Hillis (1996). «Phylogenetic inference», in Hillis, David M; C. *Moritz and B. K. Mable (editors): Molecular Systematics, 2. ed., Sunderland, MA: Sinauer Associates. ISBN 0-87893-282-8.
- Wiley, Edward O. (1981). Phylogenetics: The Theory and Practice of Phylogenetic Systematics. New York: Wiley Interscience. ISBN 0-471-05975-7.
Посилання
- Wiley, E. O.; Siegel-Causey, D.; Brooks, D. R.; Funk, V. A. (1991). The Compleat Cladist: a primer of phylogenetic procedures 19. Lawrence, Kansas: Museum of Natural History, University of Kansas. ISBN 0-89338-035-0. doi:10.5962/bhl.title.4069.(англ.)
- Example of cladistics used in textual criticism
- Journey into Phylogenetic Systematics
- Phylogenetics Primer з Talk.Origins
- Willi Hennig Society
- Cladistics: The International Journal of the Willi Hennig Society (ISSN 0748-3007)
- Phylogenetic inferring on the T-REX server
|
|
Ця стаття належить до добрих статей української Вікіпедії. |