Докази еволюції
До́кази еволю́ції — наукові дані і концепції, які підтверджують походження всіх живих істот на Землі від спільного предка[~ 1]. Завдяки цим доказам основи еволюційної теорії отримали визнання наукової спільноти, а провідною системою уявлень про процеси видоутворення стала синтетична теорія еволюції[1][~ 2].
| Частина серії статей на тему: |
| Еволюційна біологія |
|---|
.jpg.webp) Скам'янілість археоптерикса[⇨] |
|
|
Процеси |
|
Розвиток теорії
|
|
Дисципліни і застосування
|
|
Вплив на соціум
|
|
|
Еволюційні процеси спостерігаються як в природних, так і в лабораторних умовах. Відомі випадки утворення нових видів[2]. Описані також випадки розвитку нових властивостей завдяки випадковим мутаціям[3]. Факт еволюції на внутрішньовидовому рівні доведений експериментально, а процеси видоутворення спостерігались безпосередньо в природі.
З метою отримання відомостей про еволюційну історію життя, палеонтологи аналізують викопні рештки організмів. Ступінь спорідненості між сучасними видами можливо визначити порівнюючи їх будову[4], геноми, розвиток ембріонів (ембріогенез)[5]. Додаткове джерело інформації про еволюцію — закономірності географічного поширення тварин і рослин, які вивчає біогеографія[6]. Всі ці дані складаються в єдину картину — еволюційне дерево життя.
Видима еволюція
Зміни в популяціях, які спостерігаються сьогодні, доводять не тільки існування еволюції, а також і ряду механізмів, необхідних для еволюційного походження всіх видів від спільного предка. Встановлено, що геноми схильні до різноманітних мутацій, серед яких переміщення інтронів, дуплікація генів, рекомбінації, транспозиції, ретровірусні вставки (горизонтальний перенос генів), заміна, видалення і вставка окремих нуклеотидів, а також хромосомні перебудови. Відомі такі хромосомні перебудови як дуплікація генома (поліплоїдія), нерівний кросинговер, хромосомна інверсія, транслокація, ділення, злиття, дуплікація і видалення хромосом[7][8]. Також спостерігаються зміни будови організмів[9][10] і функціональні зміни[9][11] — різноманітні адаптації[12], поява здатності засвоювати новий вид їжі (в тому числі — нейлон і пентахлорфенол, виробництво яких розпочалось у 1930-х роках)[13] і т. д. Крім того, були виявлені різноманітні проміжні етапи виникнення нових видів, що свідчить про плавний характер видоутворення.
Еволюція кишкової палички в лабораторії

Групі біологів із Мічиганського університету під керівництвом Річарда Ленскі вдалось змоделювати в лабораторії процес еволюції живих організмів на прикладі бактерії кишкової палички (Escherichia coli).
Через велику швидкість розмноження зміна поколінь кишкової палички відбувається вкрай швидко, тому учені очікували, що тривале спостереження продемонструє механізми еволюції у дії. На першому етапі експерименту, в 1988 році, 12 колоній бактерій були поміщені в ідентичні умови: ізольоване поживне середовище, в якому було присутнє тільки одне джерело живильних речовин — глюкоза. Крім того, в середовищі був цитрат, який в присутності кисню ці бактерії не можуть використовувати як джерело їжі. За 20 років в колоніях змінилось понад 44 тисячі поколінь бактерій.
Учені спостерігали за змінами, які відбувались із бактеріями. Більшість із них мали однаковий характер у всіх популяціях — наприклад, розмір бактерій збільшувався, хоча і різними темпами. Проте приблизно між поколіннями номер 31000 і 32000 в одній з популяцій відбулися кардинальні зміни, які не спостерігалися в інших. Бактерії набули здатності засвоювати цитрат. Використовуючи збережені зразки бактерій із різних поколінь, дослідникам вдалось встановити, що початок серії змін, які привели до утворення нового різновиду бактерій, стався в межах покоління номер 20000 і тільки в цій колонії[3][14].
Досліди Григорія Шапошникова
Наприкінці 1950-х — на початку 1960-х років відомий радянський біолог Георгій Шапошников провів ряд дослідів, в процесі яких проводилась зміна кормових рослин у різних видів попелиць. Під час дослідів вперше спостерігалася репродуктивна ізоляція використаних в експерименті особин від вихідної популяції, що свідчить про утворення нового виду[15][16][17][18].
Еволюція вірусів
Частота виникнення мутацій у вірусів набагато вища, ніж у багатоклітинних організмів, тому їх еволюція відбувається в десятки разів швидше. Найбільш помітним для людей еволюційним пристосуванням вірусів є набуття стійкості проти лікарських препаратів. Коли в 80-х з'явилися перші засоби для лікування ВІЛ, у більшості пацієнтів ефективне лікування тривало не достатньо довго, оскільки вірус набував стійкості проти антиретровірусних препаратів впродовж кількох років[19], що робило лікування ними неефективним. Зараз, щоб сповільнити еволюцію вірусу ВІЛ до форм нечутливих до ліків, використовують для лікування хворих на СНІД використовують суміші препаратів з різними механізмами дії, в цьому випадку виникнення в одного з екземплярів вірусу мутації, що захищає від дії певного препарату, не гарантує цьому екземпляру вірусу виживання, що сповільнює закріплення такої мутації.
Швидка еволюція ящірок в нових умовах
Відповідно до опублікованого в 2008 році дослідження, у популяції стінних ящірок Podarcis sicula всього за 36 років (вкрай короткий для еволюції період) змінилися розмір і форма голови, збільшилася сила укусу і розвинулися нові структури в травному тракті. Ці фізичні зміни відбулися одночасно з серйозними змінами в популяційній щільності і соціальній структурі.
У 1971 році п'ять пар дорослих особин Podarcis sicula з острова Под Копісте були перевезені на сусідній острів Под Маркару (ці хорватські острови розташовані на півдні Адріатичного моря, недалеко від Ластово). Умови на новому місці були близькі до звичного середовища, наземні хижаки були відсутні, і за три десятиліття ящірки природним чином поширилися на острові (витіснивши при цьому місцеву популяцію Podarcis melisellensis). Коли дослідники повернулися на Под Маркару, вони виявили, що ящірки на цьому острові істотно відрізняються від ящірок на Под Копісте. Порівняння мітохондріальної ДНК показало, що популяції генетично не відрізняються, але в той же час в популяції з Под Маркару середній розмір особин збільшений, задні кінцівки коротше, максимальна швидкість бігу менше, а також відрізняється реакція на напад хижаків. Ці зміни будови і поведінки пояснюються меншою активністю хижаків на Под Маркару і наявністю щільнішої рослинності, що сприяло маскуванню.
Крім того, у нової популяції істотно змінилися розмір і форма голови, вона стала масивнішою, збільшилася сила укусу. Ці фізичні зміни відповідають зміни раціону: P. sicula з Под Копісте переважно комахоїдні, а дієта ящірок на Под Маркару в залежності від сезону майже на дві третини складається з рослинної їжі. Рослини на новому місці проживання жорсткі і волокнисті, а потужні щелепи дозволяють якісніше відкушувати і подрібнювати рослинну їжу.
Найбільш неочікуваною[20] відмінністю виявилась наявність у нової популяції у кишечнику ілеоцекальних клапанів, які призначені для сповільнення проходу їжі за рахунок створення камер бродіння, в яких мікроби можуть розбивати важко перетравлювані шматочки рослинності. Ці клапани ніколи раніше не були виявлені у цього різновиду ящірок, включаючи початкову популяцію з острова Под Копісте. Фахівці називають їх «еволюційним нововведенням, новою властивістю, яка була відсутня у предкової популяції і раніше в ході еволюції у цих ящірок».[21] Крім того, в кишечнику нової популяції присутні нематоди, які відсутні у вихідної популяції.
Зміни в дієті також викликали зміни в щільності популяції і соціальній структурі. Через те, що рослини — це набагато більше, передбачуване і надійніше джерело їжі, ніж комахи, кількість ящірок збільшилася. Крім того, пропала необхідність ганятися за здобиччю, а це призвело до того, що ящірки перестали агресивно захищати свої території.[22]
У той саме час, деякі вчені не поспішають із висновками. Так, наприклад, на думку Ендрю Гендрі, професора біології з Університету Макгілла у Монреалі, ще варто обговорювати те, як інтерпретувати подібні зміни, — чи мають вони генетичну основу, чи це є Модифікаційною мінливістю у відповідь на вплив навколишнього середовища. Втім, він додає, що такого роду зміни дійсно драматичні, тому було б логічним кроком підтвердити генетичну основу таких змін[23].
Розвиток стійкості до антибіотиків і пестицидів
Розвиток і поширення стійкості до антибіотиків у бактерій і стійкості до пестицидів у рослин та комах є доказом еволюції видів. Поява стійких до ванкоміцину форм золотистого стафілококу та небезпечність, яку вони становлять для пацієнтів лікарень, — це прямий результат еволюції шляхом природного відбору. Ще один приклад — розвиток штамів шигели, стійких до антибіотиків із групи сульфаніламідів[24]. Поява малярійних комарів, стійких до ДДТ, і розвиток у австралійських популяцій кролів стійкості до міксоматозу демонструють дію еволюції в умовах тиску відбору при швидкій зміні поколінь.
За останні 50 років зареєстровано понад 2500 випадків адаптації комах-шкідників до різних пестицидів. Тому для боротьби зі шкідниками замість звичайних отрут намагаються використовувати біологічні пестициди — ентомопатогенні віруси. Зокрема для боротьби з яблуневою плодожеркою Cydia pomonella (личинки якої — це ті самі «черв'яки» в червивих яблуках) активно використовується Cydia pomonella грануровірус (CpGV). Але у 2003 році були зафіксовані перші випадки зниження ефективності вірусу, і до 2005 року з'явилися популяції яблуневої плодожерки у яких стійкість до вірусу зросла у 100 тис. разів у порівнянні з початковою. Встановлено, що причиною стійкості є мутація в статевій Z-хромосомі[25].
Приклади видоутворення
Згідно із синтетичною теорією еволюції, утворення нових видів відбувається внаслідок поділу особин одного виду на групи, які не схрещуються між собою. Такий поділ може бути викликаний різними причинами, такими як наявність непереборних географічних бар'єрів (алопатричне видоутворення) чи розділення по екологічних нішах всередині одного ареалу (симпатричне видоутворення). Після виникнення між популяціями різноманітних типів ізоляцій, популяцій починають еволюціонувати незалежно, внаслідок чого між популяціями поступово накопичуються генетичні відмінності, і через деякий час досягається генетична несумісність — схрещування виявляється неможливим.
Крім того, нові види можуть виникати практично миттєво (в геологічному масштабі часу) шляхом поліплоїдії (кратного збільшення кількості хромосом в клітині) і гібридизації (схрещення різних видів).
Спостереження за сучасними видами показують, що видоутворення безперервно відбувається в існуючих популяціях.
Проміжні етапи видоутворення

У визначенні поняття біологічного виду істотне значення має практично повна репродуктивна ізоляція в природних умовах[26]. Якщо видоутворення відбувається поступово, як наслідок незначних послідовних змін, тоді і сьогодні ми маємо спостерігати всі можливі стадії видоутворення, а це означає і всі можливі стадії репродуктивної ізоляції: вільне схрещування між популяціями, ускладнене схрещування, зниження плодючості потомства, безплідне потомство і повну генетичну ізоляцію. Якщо нічого подібного серед сучасних видів виявити не вдалося б, то тим самим основи еволюційної теорії були б спростовані[27].
Існує безліч прикладів того, як різні види можуть схрещуватися за виняткових умов. Наприклад, у чорної ворони і сірої ворони різні ареали, але на перетині цих ареалів види можуть утворювати гібриди. Ще один приклад — різні види риб роду чукучанів з ріки Платт, які живуть поруч, але рідко схрещуються[28].
Неповне видоутворення можна спостерігати у різних кільцевих видів[29]: представники виду живуть навколо гір, водойм і т. п., при цьому сусідні популяції схрещуються між собою і незначно відрізняються, а крайні форми розрізняються значно і не схрещуються (див. діаграму). Приклад кільцевого виду — безлегенева саламандра (Ensatina eschscholtzii), що живе на схилах гір поблизу тихоокеанського узбережжя Північної Америки. Залежно від району проживання навколо гір саламандри утворюють різні форми, поступово змінюють свої морфологічні та екологічні характеристики. Крайні форми Ensatina eschscholtzii eschscholtzii і Ensatina eschscholtzii klauberi, перша з яких має червонувате забарвлення, а друга чорно-біле, співіснують у вузькій зоні гір на півдні Каліфорнії, проте між собою не схрещуються[29][30]. Інші приклади колових видів — зелений вівчарик (Phylloscopus trochiloides), оленяча миша (Peromyscus maniculatus), наземний равлик (Partula olympia), дрозофіла (Drosophila paulistorum).
У багатьох гібридів знижена фертильність. У гібридів землерийок із роду бурозубок розмноження ускладнене внаслідок різниці хромосомних наборів. Той же ефект спостерігався в лабораторії при схрещуванні дрозофіл Drosophila pseudoobscura із Юти і Каліфорнії. Фертильність також знижена у гібридів жаб з родини бомбінових кумки червоночеревої і кумки жовточеревої[31].
Багато інших видів можуть давати безплідне гібридне потомство. Серед них — різноманітні земноводні (наприклад — деякі види жаб із роду Rana) і ссавці (наприклад, мул — гібрид від схрещування коня і віслюка — як правило, безплідний). Гібриди гребінчастого тритона і мармурового тритона безплідні внаслідок наявності непарних хромосом[32].
При схрещуванні ряду інших видів досягається запліднення, але пізніше зародок гине. Як приклад можна навести леопардову жабу (Rana pipiens) і лісову жабу (Rana sylvatica) із роду справжніх жаб[33]. Аналогічна ситуація спостерігається у дрозофіл і у деяких рослин, таких як бавовник звичайний (Gossypium hirsutum) і бавовник барбадоський (Gossypium barbadense)[34].
- Білий ведмідь
Конкретним прикладом масштабних еволюційних змін при неповному видоутворенні є білий ведмідь, який, незважаючи на спорідненість із бурим ведмедем (Ursus arctos), очевидну через той факт, що ці види можуть схрещуватися і давати плодюче потомство[35], набув значних фізіологічних відмінностей з бурим ведмедем. Ці відмінності дозволяють білому ведмедю комфортно жити в умовах, в яких бурий ведмідь не вижив би. Зокрема, білий ведмідь здатен проплисти десятки кілометрів у крижаній воді, його забарвлення зливається зі снігом, він не замерзає в Арктиці. Все це можливо завдяки конкретним змінам: біле забарвлення сприяє маскуванню хижака при полюванні на тюленів; пустотілі волоски збільшують плавучість і зберігають тепло; шар підшкірного жиру, товщина якого до зими доходить до 10 сантиметрів, забезпечує додаткову теплоізоляцію; подовжена, в порівнянні з іншими ведмедями, шия дозволяє легше тримати голову над водою під час плавання; збільшені лапи з перетинками діють як весла; невеликі горбки і порожнини-присоски на підошвах зменшують небезпеку посковзнутися на льоду, а щільна шерсть на підошвах захищає лапи від сильного холоду і забезпечує тертя; вуха менші, ніж у інших ведмедів, і зменшують втрати тепла; повіки діють як сонячні окуляри; його зуби гостріші, ніж у інших ведмедів, і більше підходять для повністю м'ясного раціону; збільшений об'єм шлунка дозволяє голодному хижакові з'їсти відразу цілого тюленя, крім того, білий ведмідь здатен обійтися без їжі до дев'яти місяців за рахунок переробки сечовини[36][37].
Спостережуване видоутворення
Судячи з палеонтологічного літопису і з вимірювань швидкості мутацій, повна несумісність геномів, яка робить неможливим схрещування, досягається в природі в середньому за 3 млн років[38]. Це означає, що спостереження за утворенням нового виду в природних умовах в принципі є можливим, але це — рідкісна подія. Водночас, за лабораторних умов швидкість еволюційних змін може бути збільшеною, тому є підстави сподіватися на спостереження видоутворення у лабораторних тварин[39][40].
Відомо багато випадків видоутворення через гібридизацію і поліплоїдизацію у таких рослин, як коноплі, кропива, первоцвіт, редька, капуста, а також у різних видів папороті. У ряді випадків видоутворення у рослин відбувалося без гібридизації та поліплоїдизації (кукурудза[41], Stephanomeria malheurensis із родини айстрових[42]).
Дрозофіли, також відомі як плодові мушки, належать до найбільш вивчених організмів. З 1970-х років зафіксовано багато випадків видоутворення у дрозофіл. Видоутворення відбувалося, зокрема, за рахунок просторового розділення, розділення за екологічними нішами в одному ареалі, зміни поведінки в паруванні, дизруптивного відбору, а також за рахунок поєднання ефекту засновника з ефектом пляшкового горла.
Видоутворення спостерігалося в лабораторних популяціях кімнатних мух, мухи-осетниці Eurosta solidaginis, яблуневої плодової мухи Rhagoletis pomonella, борошняних жуків, комарів та інших комах.
Відомі випадки, коли в результаті тиску відбору (у присутності хижаків) одноклітинні зелені водорості з роду хлорела утворювали багатоклітинні колоніальні організми, а бактерії за аналогічних умов змінювали будову і збільшували розміри (з 1,5 до 20 мікрометрів за 8-10 тижнів). Чи є ці випадки прикладами видоутворення, залежить від того, яке визначення виду використовується (за безстатевого розмноження неможливо долучити критерій репродуктивної ізоляції)[43][44].
Видоутворення також спостерігалося і у ссавців. Шість випадків видоутворення у хатніх мишей на острові Мадейра за останні 500 років були наслідком виключно географічної ізоляції, генетичного дрейфу і злиття хромосом. Злиття двох хромосом — це найпомітніша відмінність геномів людини і шимпанзе, а у деяких популяцій мишей на Мадейрі за 500 років було дев'ять подібних злить[2][45].
- Яблунева плодова муха
Яблунева плодова мухаRhagoletis pomonella є прикладом спостережуваного симпатричного видоутворення (тобто видоутворення в результаті поділу за екологічними нішами)[46][47]. Спочатку вид мешкав у східній частині США. До появи європейців личинки цих мух розвивалися тільки в плодах глоду. Однак із завезенням в Америку яблунь (перша згадка яблунь в Америці — 1647 рік), відкрилася нова екологічна ніша. В 1864 році личинок Rhagoletis pomonella було виявлено в яблуках, і тим самим зафіксовано яблуневу расу цього виду[48]. За півтора століття спостережень раси дуже сильно розійшлися. Вони майже не схрещуються одна з одною (рівень гібридизації не перевищує 4-6 %). Яблунева раса парується майже виключно на яблунях, а глодова — на глоді, що, враховуючи різний час дозрівання плодів, призводить до репродуктивної ізоляції. Незабаром є можливим перетворення цих рас в самостійні види.
У плодових мух, або ж мух-осетниць відомо ще кілька видів-двійників, які живуть на різних видах рослин, — імовірно, видоутворення у них відбувалось саме за описаною схемою[49][50][51].
Еволюційне дерево
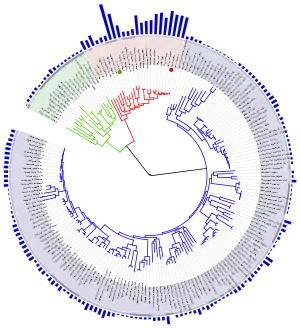
Загальноприйнята в науковій спільноті класифікація живих організмів являє собою багаторівневу ієрархічну структуру: організми поділяються на царства, царства поділяються на типи, типи — на класи, класи — на ряди, і так далі. Внаслідок такого розгалуження утворюється філогенетичне дерево[~ 3]. Існують суперечності між біологами з приводу віднесення окремих видів до конкретних груп, але ці суперечності мають поодинокий характер. На практиці біологічні класифікації, побудовані на основі різних ознак, в тенденції наближаються до однієї і тієї ж деревоподібної ієрархічної схеми — природної класифікації. А значить, можливо, в основі цієї класифікації існує закономірність.
Саме такий результат можна очікувати при евлюційному походженні тварин від спільного предка[~ 4]. Розгалуження філогенетичного дерева відповідає поділу популяцій в процесі видоутворення.
Як правило, об'єкти, що виникли не в ході еволюції, не мають таких властивостей. Неможливо об'єктивно побудувати ієрархію елементарних частинок, хімічних елементів, планет Сонячної системи. Також не існує об'єктивної ієрархії таких свідомо створених об'єктів як книги в бібліотеці, будинки, меблі, машини. Можна при бажанні об'єднати ці об'єкти в різноманітні ієрархії, але не існує єдиної об'єктивної ієрархії, яка принципово краща, ніж всі інші[52].
Існує низка статистичних методів для точної оцінки того, наскільки об'єкти із різними якостями вкладаються в ту чи іншу ієрархію[53]. Ці методи вимірюють так званий «філогенетичний сигнал», дозволяючи відрізнити уявні закономірності від об'єктивних. Наприклад, у будь-якого «генеалогічного древа» автомобілів буде низький рівень філогенетичного сигналу[54][55]. У еволюційного дерева і у різних його частин, навпаки, стабільно чіткий філогенетичний сигнал[55][56][57].
Є декілька джерел даних, на основі який можна робити висновки про ступінь спорідненості між видами. Якщо існує єдине еволюційне дерево, яке об'єднує всі види в об'єктивну генеалогію, то всі дані повинні підтверджувати цю генеалогію[58]. Філогенетичні дерева, побудовані незалежно, повинні відповідати один одному. Найбільш простий, хоча не дуже точний спосіб побудувати таке дерево — порівняння будови організмів тварин: чим менше розбіжностей між видами, тим менше поколінь відділяють їх від спільного предка. Палеонтологічний літопис дозволяє встановити спорідненість між класами тварин. Наприклад, знайдені рештки пернатих динозаврів свідчать про спорідненість між рептиліями і птахами. Біогеографія і ембріологія також дають інформацію про еволюційну близькість видів. Найточніше джерело даних, яке було недоступним за часів Дарвіна — порівняльний аналіз геномів різних організмів. Еволюційне дерево можна побудувати по кожному окремому гену, дослідники також розглядають різноманітні послідовності, які не кодують білки́.
Всі ці джерела інформації дають однакову картину з точністю до похибки використовуваних методів. Той факт, що еволюційні дерева, які побудовані за різними даними, відповідають один одному, пояснюється еволюційною теорією. Інші пояснення відсутні: наприклад, немає ніяких причин, чому організми, подібні за будовою, повинні мати подібні некодуючі послідовності в геномі, якщо вони не походять від одного предка[59].
Палеонтологічні докази
| 1850 |  |
| 1950 | 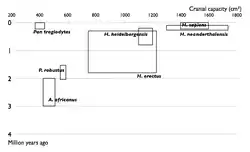 |
| 2002 | 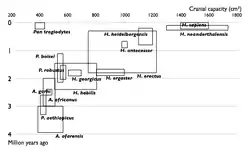 |
| Прогрес у вивченні еволюції людини. Збільшення числа відомих видів гомінін у часі. кожен вид зображений у вигляди прямокутника, який показує межі, в який в яких варіював обсяг черепа, і місце виду в палеонтологічному літописі. Видно поступове заповнення проміжків між видами. | |
Як правило, рештки рослин і тварин розкладаються й зникають без сліду. Але іноді біологічні тканини заміщуються мінеральними речовинами, і утворюються скам'янілості. Зазвичай знаходять скам'янілі кістки або раковини, тобто тверді частини живих організмів. Іноді знаходять відбитки слідів тварин або сліди їх життєдіяльності. Ще рідше знаходять тварину цілком — вимороженою в льоду в районах сучасної вічної мерзлоти, або яка потрапила в закам'янілу пізніше смолу прадавніх рослин — бурштин, або в іншу природну смолу — асфальт.
Вивченням викопних рештків займається палеонтологія. Зазвичай, осадові породи залягають шарами, тому більш глибокі шари містять скам'янілості з більш раннього періоду (принцип суперпозиції в геології). А отже, порівнюючи викопні форми із послідовних нашарувань, можна робити висновки про основні напрямки еволюції живих організмів. Для оцінки віку скам'янілостей використовуються численні методи геохронології.
При погляді на палеонтологічний літопис можна зробити висновок, що життя на Землі істотно змінювалось. Чим глибше в минуле ми дивимося, тим менше бачимо спільного з сучасною біосферою. Перші прокаріоти (найпростіші одноклітинні, що не володіють оформленим клітинним ядром) з'являються приблизно 3,5 млрд років тому. Перші одноклітинні еукаріоти з'являються 2,7-1,75 млрд років тому. Приблизно через мільярд років, 840 млн років тому, в палеонтологічному літописі з'являються перші багатоклітинні тварини, представники хайнаньської фауни. Відповідно до опублікованого в 2009 році дослідження, ймовірно, понад 635 млн років тому вже існували багатоклітинні, що відносяться до одного з сучасних типів — губки[60]. В період «кембрийського вибуху», 540—530 млн років тому, за геологічно короткий проміжок часу, в геологічному літописі з'являються рештки представників більшості сучасних типів, що мають скелети, а ще через 15 млн років — перші примітивні хребетні, схожі на сучасних міногів[61]. Щелепні риби з'являються 410 млн років тому, комахи — 400 млн років тому, і ще 100 млн років на суходолі панують папоротеподібні, а основними групами наземної фауни залишаються комахи і земноводні. З 250 по 65 млн років тому на Землі панівне положення «верховних хижаків» і великих травоїдних займали динозаври та інші плазуни, найпоширенішими рослинами були саговники і інші групи голонасінних. Перші викопні рештки квіткових з'являються 140—130 млн років тому, а початок їх широкого розповсюдження відноситься до середини крейдового періоду (близько 100 млн років тому). Ця картина відповідає походженню всіх видів від одноклітинних організмів і не має інших наукових пояснень[62].
Відомий доказ еволюції — наявність так званих перехідних форм, тобто організмів, що поєднують в собі характерні ознаки різних видів.[~ 5] Як правило, говорячи про проміжні (або «перехідні») форми мають на увазі викопні види, хоча проміжні види не завжди вимирають. На основі філогенетичного дерева теорія еволюції пророкує, які проміжні форми можуть бути знайдені, а які — ні. Відповідно до наукових методів, прогнозування, які справджуються, підтверджують теорію. Наприклад, знаючи будову організмів плазунів і птахів, можна передбачити деякі особливості перехідної форми між ними. Можна прогнозувати можливість знайти рештки тварин, подібних рептиліям, але з пір'ям, або рештки тварин, подібних птахам, але з зубами чи з довгими хвостами зі скелетом з незріщених хребців. При цьому можна передбачити, що не будуть знайдені перехідні форми між птахами і ссавцями, наприклад — викопні ссавці з пір'ям або подібні птахам викопні з кістками середнього вуха як у ссавців[63].
Невдовзі після публікації «Походження видів» були виявлені рештки археоптерикса — проміжної форми між рептиліями і птахами. Для археоптерикса властиво диференційоване оперення (типова риса птахів), а по будові хребта він мало відрізнявся від динозаврів із групи компсогнатів. Довгий хвостовий відділ хребта з 20 хребців, пальці з розвинутими фалангами на передніх кінцівках, наявність черевних ребер, відсутність дзьоба і низка інших ознак характерні для рептилій. Водночас загальна форма тіла, ключиці, які зрослись, трипалі передні кінцівки і характер оперення, яке містить складно влаштовані махові пір'я, — характерні для птахів. Таким чином археоптерикс займає по морфології проміжне положення між рептиліями і птахами, тобто є перехідною формою[~ 5]. Пізніше були знайдені і інші перехідні форми між рептиліями і птахами[64].
Відомо чимало інших перехідних форм, у тому числі — від безхребетних до риб, від риб до чотириногих, від земноводних до рептилій і від рептилій до ссавців[65].
У деяких випадках викопні перехідні форми виявити не вдалося, наприклад — немає слідів еволюції шимпанзе (імовірно, це пояснюється відсутністю умов для утворення скам'янілостей в лісах, де вони живуть)[66], немає слідів війчастих червів, а цей клас об'єднує понад 3500 видів[67]. Зрозуміло, щоб заперечити спростовуваність теорії еволюції, недостатньо вказати на подібні прогалини в палеонтологічному літописі. Щоб спростувати еволюційну теорію, треба було б пред'явити скелет, який не відповідає філогенетичному дереву або не вкладається в хронологічну послідовність. Так, у відповідь на питання про те, яка знахідка могла б спростувати еволюційну теорію, Джон Голдейн відповів: «Викопні кролі в докембрії!»[68] Були знайдені мільйони скам'янілостей[69] близько 250 000 викопних видів[70], і кожна знахідка — це перевірка теорії еволюції, а пройдена перевірка підтверджує теорію.
В тих випадках, коли палеонтологічний літопис виявляється особливо повним, з'являється можливість побудувати так звані філогенетичні ряди, тобто такі ряди видів (родів і т. д.), які послідовно змінювали один одного в процесі еволюції. Найбільш відомі філогенетичні ряди людини і коня, також як приклад можна навести еволюцію китоподібних[71].
Еволюція людини
Перший екземпляр неандертальця був знайдений у 1856 році, за три роки до видання «Походження видів». На момент виходу книги не було відомо ніяких інших викопних решток, які б підтверджували еволюційне походження людини і людиноподібних мавп від спільного предка. Хоча неандерталець є незалежним видом із роду людей, з тих пір було знайдено безліч скелетів проміжних форм між давніми антропоїдами і людиною.
Оскільки спільний предок людини і шимпанзе пересувався на чотирьох кінцівках, а мозок у нього був більше ніж у шимпанзе, згідно із теорією Дарвіна в процесі еволюції повинно було розвинутися прямоходіння, а також мав збільшитися об'єм мозку. Таким чином, повинний був існувати один із трьох варіантів перехідних форм:
- Проміжний розмір мозку, розвиток прямоходіння.
- Розмір мозку приблизно такий як у шимпанзе, розвинене прямоходіння.
- Більший мозок, прямоходіння не розвинене[72].
У 1920-х роках в Африці були виявлені рештки істоти, яку Раймонд Дарт назвав австралопітеком. Пізніше були знайдені й інші рештки австралопітеків, в тому числі — відома Люсі і череп AL 444-2. Австралопітеки жили у східній та північній Африці 4-2 млн років тому. Об'єм мозку австралопітека був трохи більше, ніж у шимпанзе. Кістки таза за будовою найбільш близькі до кісток таза людини. Будова черепа характерно для прямоходячих тварин, що можна визначити зокрема за foramen magnum — отвором в потиличній кістці, що з'єднує порожнину черепа з хребетним каналом. Більше того, в Танзанії в скам'янілому вулканічному попелі були виявлені сліди, залишені двоногими гомінідами. Вік слідів становить 3,6 млн років[73][74]. Таким чином, австралопітеки є «перехідною формою номер два»: у них було розвинене часткове прямоходіння, але мозок був приблизно як у шимпанзе[75].
Пізніше були знайдені рештки ардипітека, вік яких становить 4,5 млн років. Аналіз скелету показав, що по землі ардипітеки пересувалися на двох задніх кінцівках, а по деревах лазили на всіх чотирьох[76][77]. Прямоходіння у ардипітеків було слабо розвинене в порівнянні з наступними гомінідами, вони не могли долати великі відстані[78]. Ардипітек являє собою перехідну форму між спільним предком людини і шимпанзе та австралопітеком.
Отже, в результаті еволюції у гомінід спочатку розвинулося прямоходіння, і тільки потім почалося суттєве збільшення об'єму мозку. У австралопітеків, які жили 4-2 млн років тому, об'єм мозку був близько 400 см³, приблизно як у шимпанзе. Людина уміла (Homo habilis) жила 2,4—1,4 млн років тому, розмір мозку становив у неї 500—640 см³. Людина працююча (Homo ergaster) жила 1,9—1,4 млн років тому, розмір мозку — 700—850 см³. Людина прямоходяча (Homo erectus) жила 1,4—0,2 млн років тому, розмір мозку становив від 850 см³ у ранньої особини до 1100 см³ у пізніх. Гейдельберзька людина (Homo heidelbergensis) жила 600—350 тисяч років тому, розмір мозку становив 1100—1400 см³. Неандертальці жили 350—25 тисяч років тому, розмір мозку становив 1200—1900 см³. 200 тисяч років тому з'явилася людина розумна (Homo sapiens), розмір мозку людей становить 1000—1850 см³. Згідно із сучасними уявленням, не всі перелічені види були безпосередніми предками сучасних людей. Зокрема, неандертальці розвивалися паралельно з Homo sapiens. Тим не менш, на прикладі цих видів можна спостерігати напрямок еволюції гомінідів. Ці види добре представлені в палеонтологічному літописі[79][80].
Згідно із Дарвіном, «в ряду форм, які непомітно переходять одна в іншу від будь-якої мавпоподібної істоти до людини в її сучасному стані, було б неможливо точно вказати, якій саме з цих форм слід вперше дати найменування „людини“»[81]. Саме з цією проблемою зіткнулись сучасні палеонтологи, які змушені ділити знайдені рештки гомінідів на різні роди. Наприклад, черепи KNM ER 1813, KNM ER 1470 і OH 24 («Твіггі») в різний час класифікувались як Australopithecus habilis і як Homo habilis, лише тому що вони опинилися на штучному «кордоні», довільно проведеному між практично нерозрізненими родами[82].
Еволюція коня
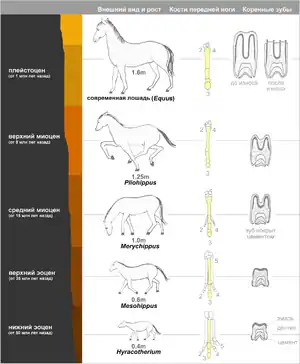
Еволюція коня достатньо повно простежується в Північній Америці. Найдавніший представник коневих — гіракотерій (Hyracotherium, також відомий як Eohippus) — був розміром з лисицю і жив у Північній Америці 54 млн років тому (нижній еоцен), а потім поширився в Європу і в Азію[84]. Це була тварина стрункої будови, з короткими ногами, але пристосоване для бігу. У нього було чотири пальці на передніх ногах і три пальці на задніх, пальці розміщувались практично вертикально. Різці були маленькі, кутні зуби приплющені і покриті зверху емаллю.
Вірогідний шлях розвитку коней від гіракотерій до сучасного виду вміщує не менше 12 родів і декілька сотень видів. При цьому відбулись наступні зміни:
- Збільшення розміру (з 0,4 до 1,6 м)
- Подовження ніг і стоп
- Редукція бічних пальців
- Подовження і потовщення середнього пальця
- Збільшення ширини різців
- Заміщення премолярів кутніми зубами
- Подовження зубів
- Збільшення коронки кутніх зубів
Судячи по викопним рослинам із різних шарів, болотисті місцевості, в яких мешкав гіракотерій, поступово ставали сухіше. Предки коней почали покладатись на покращений за рахунок піднятої голови огляд і на високу швидкість бігу для порятунку від хижаків. Якщо при ходінні по м'якому болотистому ґрунту велика кількість пальців на кінцівках була виправдана, то у змінених умовах існування для первісних коней стало вигіднішим, коли бічні пальці почали у них поступово відмирати, а середні — розвиватися. Зміна будови зубів пояснюється зміною раціону, а саме — переходом з м'якої рослинності на траву та листя.
Іноді у коней розвиваються один чи два зайвих пальця, частіше зустрічається 2-й (внутрішній) палець з цілком розвиненими п'ястковими чи плюсневими кістками, цілком розвинутими суглобами пальця і копитом, яке, проте, рідко торкається землі. У випадку розвитку двох зайвих пальців нога отримує схожість із ногами предкових видів.
Морфологічні докази
У ході еволюції кожний новий організм не проектується з нуля, а утворюється зі старого за рахунок послідовності невеликих змін. В утворених таким чином структур є ряд характерних особливостей, які вказують на їх еволюційне походження. Порівняльно-анатомічні дослідження дозволяють виявити такі особливості.
Зокрема, еволюційне походження виключає можливість цілеспрямованого запозичення вдалих конструкцій в інших організмів. Тому у різних, не споріднених видів для вирішення схожих завдань використовуються різні органи. Наприклад, крило метелика і крило птаха розвиваються з різних зародкових листків, крила птахів є видозміненими передніми кінцівками, а крила метелика — складки хітинового покриву. Подібність між цими органами поверхневе і є наслідком їх конвергентного походження. Такі органи називають аналогічними[85][86].
Протилежна ситуація спостерігається у близькоспоріднених видів: для абсолютно різних завдань використовуються схожі за будовою органи. Наприклад, передні кінцівки хребетних виконують найрізноманітніші функції, але при цьому мають загальний план будови, займають подібне положення і розвиваються з одних і тих же зачатків, тобто є гомологічними[4]. Подібність будови крила кажана і лапи крота неможливо пояснити з точки зору корисності. У той же час, теорія еволюції дає пояснення: єдину структуру кінцівки чотириногі хребетні успадкували від спільного предка[87].
Кожний вид успадковує від предкового виду більшість його властивостей — в тому числі іноді й ті, які для нового виду марні. Зміни зазвичай відбуваються за рахунок поступового послідовного перетворення ознак предкового виду. Подібність гомологічних органів, не пов'язана з умовами їх функціонування — свідчення їх розвитку в ході еволюції із спільного прототипу, наявного у предкового виду. Інші приклади еволюційних змін морфології — рудименти, атавізми, а також численні випадки специфічної недосконалості будови організмів.
Гомологічні органи
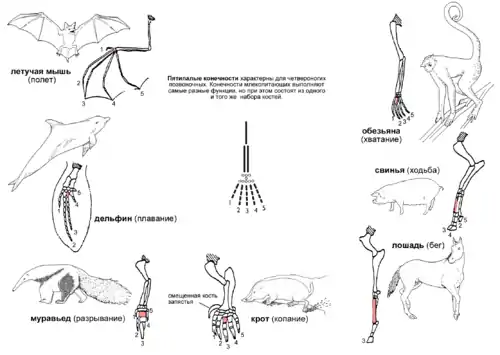
- П'ятипала кінцівка
П'ятипала кінцівка, яка характерна для чотириногих хребетних — приклад гомології органів. Більш того, простежується гомологія п'ятипалої кінцівки і плавників деяких викопних видів кистеперих риб, від яких, як видно, походять перші земноводні. Кінцівки чотириногих розрізняються за формою і пристосовані до виконання найрізноманітніших функцій у самих різних умовах. На прикладі ссавців:
- У мавп передні кінцівки витягнуті, кисті пристосовані для хапання, що полегшує лазання по деревах.
- У свині перший палець відсутній, а другий і п'ятий — зменшені. Решта два пальці довше і твердіше інших, кінцеві фаланги покриті копитами.
- У коня також замість кігтів копито, нога подовжена за рахунок кісток середнього пальця, що сприяє більшій швидкості пересування.
- Кроти мають вкорочені і потовщені пальці, що допомагає при копанні.
- Мурахоїд використовує великий середній палець для розкопування мурашників і гнізд термітів.
- У китів передні кінцівки є плавниками. При цьому число фаланг пальців збільшено в порівнянні з іншими ссавцями, а самі пальці приховані під м'якими тканинами.
- У кажана передні кінцівки перетворилися в крила за рахунок значного подовження чотирьох пальців, а гакоподібний перший палець використовується, щоб висіти на деревах.
При цьому всі ці кінцівки містять схожий набір кісток з одним і тим же відносним розташуванням[88]. Єдність структури не може бути пояснено з точки зору корисності, так як кінцівки використовуються для зовсім різних цілей.
- Частини ротового апарату комах
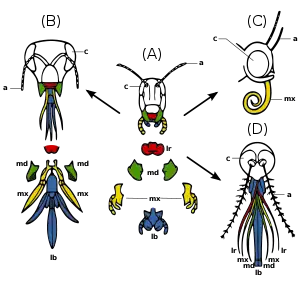
Основні частини ротового апарату комах — верхня губа, пара жвал (верхні щелепи), підглоточник (hypopharynx), дві максилли (нижні щелепи) і нижня губа (другі максилли, які зрослись). У різних видів ці складові частини розрізняються за формою і розміром, у багатьох видів деякі з частин втрачені. Особливості будови ротового апарату дозволяють комахам використовувати різні джерела їжі (див. малюнок): (A) У початковому вигляді (наприклад, у коників) сильні жвали і максилли використовуються для кусання і жування.
(B) Медоносна бджола використовує нижню губу для збору нектара, а жвалами подрібнює пилок розминає віск.
(C) У більшості метеликів верхня губа зменшена, жвали відсутні, максилли утворюють хоботок.
(D) У самиць комарів верхня губа і максилли утворюють трубку, жвали використовуються для протикання шкіри.
Аналогічні органи
Зовнішньо схожі органи та їх частини, які походять від різних початкових зачатків і мають неоднакову внутрішню будову, називаються аналогічними. Зовнішня схожість виникає в ході конвергентної еволюції, тобто в ході незалежного пристосування до подібних умов існування.
Приклади:
- Крила птахів — видозмінені передні кінцівки, крила комах — складки хітинового покрову.
- Зябра риб — утворення, пов'язані із внутрішнім скелетом, зябра багатьох ракоподібних — відрости кінцівок, ктенідіальні зябра молюсків розвиваються в мантійній порожнині, а зябра голозябрових молюсків — нарости покровів спинної сторони тіла.
- Обтічна форма тіла у водних ссавців — китів, дельфінів — і у риб.
- Колючки барбарису, кактуса — видозмінене листя, колючки глоду розвиваються із пагонів.
- Вусики винограду (які утворюються із пагонів) і вусики гороху (видозмінене листя).
- Форма різних сукулентів (рослин, які мають спеціальні тканини для запасу води), таких як кактуси і молочай[~ 6].
Повна відсутність цілеспрямованого запозичення вдалих конструкцій відрізняє еволюцію від свідомого проектування. Наприклад, пір'я — це вдала конструкція, яка допомагає у польоті, але у ссавців (в тому числі у кажанів) пір'я відсутнє. Зябра надзвичайно корисні для водних тварин, але у ссавців (таких як китоподібні) вони відсутні. Для того, щоб спростувати теорію еволюції, достатньо виявити пір'я чи зябра у якого-небудь виду ссавців[86][89].
Рудименти
Рудиментами називаються органи, які втратили своє основне призначення в процесі еволюційного розвитку організму. Рудименти також можна визначити незалежно від еволюційної теорії як редуковані структури, які мають менші можливості в порівнянні з відповідними структурами в інших організмах. Якщо рудимент і виявляється функціональним, то він виконує відносно прості або незначні функції за допомогою структур, очевидно призначених для складніших цілей. Хоча багато з рудиментарних органів абсолютно не функціональні, відсутність функцій не є необхідною умовою для рудиментарності[90]. Такі органи вкрай поширені в природі[91].
Наприклад, крило птаха — вкрай складна анатомічна структура, спеціально пристосована для активного польоту, але крила страусів не використовуються для польоту. Ці рудиментарні крила можуть використовуватися для порівняно простих завдань, таких як підтримка рівноваги на бігу і привернення уваги самиць. Таким чином, специфічна складність крил страуса неадекватна простоті завдань, для яких ці крила використовуються, і саме тому ці крила називають рудиментами. У числі інших нелітаючих птахів з рудиментарними крилами — галапагоські баклани, ківі і какапо. Для порівняння, крило пінгвіна має велике значення, діючи як плавник, а отже не може вважатися рудиментом[92].

Приклади рудиментів:
- Очі у деяких риючих і печерних тварин, таких як протей, сліпак, кріт, астіанакс мексиканський (Astyanax mexicanus, сліпа печерна риба). Часто очі приховані під шкірою.[93]
- Мала гомілкова кістка у птахів.
- Залишки волосяного покрову і тазових кісток у деяких китоподібних[94].
- У деяких змій, у тому числі у пітона, є кістки задніх кінцівок[95][96]. Ці кістки не кріпляться до хребта і відносно вільно переміщаються в черевній порожнині.
- У багатьох видів жуків, таких як Apterocyclus honoluluensis, крила лежать під надкриллями, які зрослися[97].
- У людини до рудиментів зокрема відносяться хвостові хребці, волосяний покрив тіла, вушні м'язи, бугор вушної раковини, морганієві шлуночки гортані.
Червоподібний відросток сліпої кишки (апендикс) у деяких травоїдних тварин використовується для перетравлення рослинної їжі і має велику довжину. Наприклад, у коали довжина апендикса становить від 1 до 2 метрів. Апендикс людини має довжину від 2 до 20 сантиметрів і не бере участі в розщеплюванні їжі. Всупереч поширеному переконанню, наявність у апендикса другорядних функцій не означає, що він не є рудиментом.
Теорія еволюції прогнозує які рудиментарні органи можуть бути знайдені, а які — принципово неможливі. Будь-який рудиментарний орган організму був повністю розвинений у його предків. Тому кожен виявлений рудимент повинен відповідати еволюційному дереву, інакше основи теорії еволюції довелося б переглядати. Це черговий приклад спростовуваності еволюційної теорії, тобто свідоцтво відповідності критеріям Поппера для визнання теорії науковою[98].
Атавізми
Атавізмом називається поява в особини ознак, властивих віддаленим предкам, але відсутніх у найближчих предків. Поява атавізмів пояснюється тим, що гени, які відповідають за певну ознаку, збереглись в ДНК, але в нормі не формують структури, типові для предків.
Приклади атавізмів:
- Хвостоподібний придаток у людини;
- Суцільний волосяний покрив на тілі людини;
- Додаткові пари молочних залоз;
- Задні ноги у китів;
- Задні плавники у дельфінів[99];
- Задні ноги у змій[100];
- Додаткові пальці у коней[101];
- Відновлення статевого розмноження у нечуйвітра волохатенького і у кліщів родини Crotoniidae[102][103];
Аргументи на користь еволюції аналогічні аргументам для рудиментів[104].
Недосконалість будови організмів
У результаті еволюції кожна нова конструкція утворюється зі старої за рахунок послідовності пристосовних змін. Ця особливість є причиною специфічних непорозумінь в будові живих організмів[105].
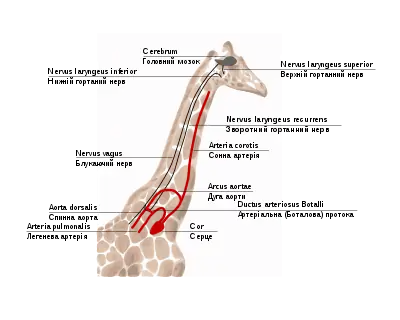
Наприклад, зворотний гортанний нерв у ссавців проходить (у складі блукаючого нерва) від мозку до серця, огинає (вже як самостійний нерв) дугу аорти і повертається до гортані. Внаслідок цього нерв проходить набагато довший шлях, ніж необхідно, а аневризма аорти може призводити до паралічу лівої голосової зв'язки. Особливо наочно проблема помітна на прикладі жирафа, у якого довжина зворотного нерва може досягати 4 метрів, хоча відстань від мозку до гортані — всього кілька сантиметрів. Таке розташування нервів і судин ссавці успадкували від риб, у яких шия відсутня[106].
Травна система ссавців перетинається з дихальними шляхами, в результаті ми не можемо одночасно дихати і ковтати, а крім того є небезпека вдавитися. Еволюційне пояснення даної обставини полягає в тому, що предками ссавців є кистепері риби (Crossopterygii) з групи Osteolepiformes, які заковтували повітря, щоб дихати.
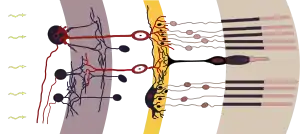
Ще один приклад — сітківка хребетних і сліпа пляма. У хребетних нервові волокна і гангліозні клітини сітківки розташовані поверх світлочутливих клітин, і світло має пройти через декілька шарів клітин, перш ніж потрапить на палички і колбочки. Сліпа пляма — це ділянка сітківки, яка позбавлена фоторецепторів, від якої відходить до мозку зоровий нерв. Для вирішення численних проблем, викликаних таким влаштуванням сітківки, у хребетних є ряд адаптацій. Зокрема, у нервових волокон, що йдуть поверх сітківки, відсутня мієлінова оболонка, що підвищує їх прозорість, але знижує швидкість передачі сигналу. Подібні за будовою ока головоногі не мають цього недоліку[107]. Це наочно показує, що і у хребетних, можливо, могли б в ході еволюції сформуватися очі без сліпих плям[108][109]. Вважається, що така будова ока пов'язана зі способом його розвитку в онто- і філогенезі. Ймовірно, у спільних віддалених предків хордових фоторецептори знаходились на спинній поверхні тіла. Потім, при формуванні нервової трубки, вони опинилися на її внутрішній поверхні, тобто на стінці нервового каналу (як у ланцетника). Очі сучасних хребетних утворюються як випинання стінок нервової трубки (очні міхури), а фоторецептори як і раніше формуються в їхній внутрішній стінці.
Такі часті патології як випадіння матки, часте запалення носоглотки, болі в попереку у людей частково пов'язані з тим, що ми використовуємо для прямоходіння тіло, сформоване еволюцією протягом сотень мільйонів років для пересування на чотирьох кінцівках[110].
Ембріологічні докази
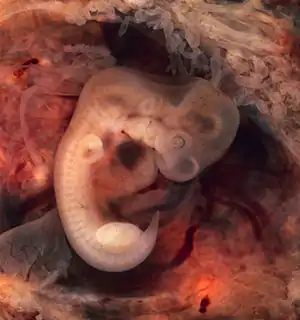
У всіх хребетних тварин спостерігається значна схожість ембріонів на ранніх стадіях розвитку: у них схожа форма тіла, є зачатки зябрових дуг і зябрових щілин, хорди, наявний хвіст, одне коло кровообігу і т. д. (закон зародкової подібності Карла Бера)[~ 7]. Проте в міру розвитку подібність між зародками різних систематичних груп поступово зменшується, і починають переважати риси, притаманні їх класам, рядам, родинам, родам і видам.
Еволюційні зміни можуть торкатися всіх фаз онтогенезу, тобто можуть призводити до змін не тільки дорослих організмів, але й ембріонів, навіть на перших етапах розвитку. Тим не менш, більш ранні фази розвитку повинні відрізнятися більшим консерватизмом, ніж пізніші, оскільки зміни на пізніших етапах, в свою чергу, мають призвести до більших змін в процесі подальшого розвитку. Наприклад, зміна типу дроблення викличе зміни в процесі гаструляції, так само як і на всіх наступних стадіях. Тому зміни, які проявилися на ранніх етапах, набагато частіше виявляються летальними, ніж зміни, що стосуються пізніших періодів онтогенезу.
Таким чином, ранні стадії розвитку змінюються відносно рідко, а значить, вивчаючи ембріони різних видів, можна робити висновки про ступінь еволюційної спорідненості[111].
У 1837 році ембріолог Карл Райхерт з'ясував, з яких зародкових структур розвиваються квадратна і зчленівна кістки в щелепі рептилій. У зародків ссавців виявлені ті ж структури, але вони розвиваються в молоточок і ковадло середнього вуха[112]. Палеонтологічний літопис підтверджує походження частин вух ссавців із кісток щелепи рептилій.
Є багато інших прикладів того, як еволюційна історія організму проявляється в ході його розвитку. У ембріонів ссавців на ранніх стадіях є зяброві мішки (Pharyngeal pouch), які не відрізняються за будовою від зябрових мішків у водних хребетних[113]. Це пояснюються тим, що предки ссавців жили у воді і дихали зябрами. Зрозуміло, зяброві мішки ембріонів ссавців в ході розвитку перетворюються не в зябра, а в структури, що виникли в ході еволюції з зябрових щілин або стінок зябрових кишень, такі як євстахієві труби, середнє вухо, мигдалики, паращитоподібні залози і тимус[114].
У ембріонів багатьох видів змій і безногих ящірок (наприклад, веретільниці ламкої) розвиваються зачатки кінцівок, але потім вони розсмоктуються[115]. Аналогічно, у китів, дельфінів і морських свиней[~ 8] немає задніх кінцівок, але у ембріонів китоподібних починають рости задні ноги, розвиваються кістки, нерви, судини, а потім всі ці тканини розсмоктуються[116].
Дарвін як приклад наводив наявність зубів у ембріонів вусатих китів[97].
У людських ембріонів на ранніх етапах розвитку (4—5 тижнів) є помітний хвіст з 10—12 хвостовими хребцями, довжина якого становить понад 10 % довжини ембріона[117][118][119]. Розвиваються не тільки хребці, а й спинний мозок, і ряд інших структур. До восьмого тижня шість крайніх хребців відмирають в результаті загибелі клітин за механізмом апоптозу. 5-й і 4-й хвостові хребці зменшуються, інші структури також відмирають і зменшуються. Макрофаги утилізують клітини, що руйнуються[117][119][120][121]. У дорослих людей залишається 4 хвостові хребці, які утворюють куприк[5][117][120][122][123].
Молекулярно-генетичні та біохімічні докази
Дарвін видав «Походження видів» майже за 100 років до розшифрування структури ДНК. Нові знання, отримані з того часу, могли б остаточно спростувати еволюційну теорію, якби вона була хибною. Натомість, аналіз ДНК дає підтвердження теорії еволюції[124]. Наявність спадкової мінливості є необхідною для еволюції, і якби виявилось, що ДНК стійка до змін, це означало би кінець теорії.[8] Але ДНК постійно мутує, до того ж ці мутації відповідають різниці між геномами різних видів. Наприклад, основні відмінності геному людини від геному шимпанзе містять 35 млн замін окремих нуклеотидів, 5 млн делецій (видалень) і інсерцій (вставок), злиття двох хромосом і дев'ять хромосомних інверсій.[125] Всі ці мутації спостерігаються і сьогодні, у протилежному випадку версію про еволюційне походження від спільного предка довелось би переглядати.
Різниця між геномами видів повинна відповідати не тільки мутаціям, які спостерігаються, але і філогенетичному дереву і палеонтологічному літопису. Подібно тому, як аналіз ДНК спроможний встановити ступіть споріднення між двома людьми, порівняння геномів дозволяє з'ясувати ступінь споріднення між видами, а знаючи кількість накопичених відмінностей, дослідники визначають час розходження двох видів, тобто час, коли існував їх останній спільний предок. Наприклад, згідно з даними палеонтології, спільний предок людини і шимпанзе жив приблизно 6 млн років тому.[126] Для того, щоб отримати спостережуване число відмінностей між геномами, на кожен мільярд нуклеотидів мало припадати в середньому 20 змін за одне покоління.[127] Сьогодні у людей швидкість мутацій становить 10—50 змін на кожен мільярд нуклеотидів за одне покоління[128], тобто дані палеонтології узгоджуються з результатами аналізу ДНК, в суворій відповідності до теорії еволюції.[129]
Біохімічна єдність життя
Носієм спадкової інформації у всіх клітинах є молекули ДНК, у всіх відомих організмів і основі розмноження — реплікація цієї молекули. В ДНК всіх організмів використовується 4 нуклеотиди (аденін, гуанін, тимін, цитозин), хоча в природі зустрічаються не менше 102 різних нуклеотидів.[130] Крім того, в природі зустрічаються 390 різних амінокислот[131], але білки всіх організмів складаються із одного і того ж набору, в якому лише 22 амінокислоти.[~ 9] При цьому можливі 1,4×1070 різноманітних інформаційно еквівалентних генетичних кодів, які використовують одні й ті самі кодони і амінокислоти.[132]
Якщо не враховувати еволюційне походження всіх організмів від спільного предка, то ніщо не заважає кожному виду мати власний генетичний код. Такий стан речей був би вкрай вигідним, тому що при цьому унеможливлювалося б подолання вірусами міжвидових бар'єрів. Проте, нічого подібного не спостерігається, і теорія еволюції виключає таку можливість: зміни генетичного коду призводять до зміни більшості білків організму, така мутація практично завжди виявляється летальної, тому код не міг значно змінитися з часів останнього спільного предка, що гарантує його універсальність.
Учені, які зробили значний внесок у виявлення структури генетичного коду в 1950-х і 1960-х роках, у своїх дослідженнях активно використовували припущення, що код є практично універсальним. Френсіс Крік, Сідні Бреннер, Джордж Гамов і інші зробили це припущення виходячи з версії про еволюційний походження від спільного предка[133], не маючи жодних прямих доказів універсальності коду[134]. Покладаючись на універсальність коду, Бреннер в 1957 році дійшов висновку про неперекривність коду (один і той же нуклеотид не може входити одночасно до складу двох або більше триплетів)[135]. Робота мала велике значення, оскільки до неї більшість дослідників вважали, що код перекривається.
У 1961 році, за п'ять років до встановлення структури генетичного коду, Френсіс Крік посилався на роботу Бреннера у статті «Спільна природа генетичного коду для білків»[136]. Спираючись на еволюційне передбачення про універсальність коду (зокрема — на тому, що у бактерій, тютюну і людей код влаштований однаково), Крік встановив такі найважливіші властивості генетичного коду як триплетність (значущою одиницею коду є поєднання трьох нуклеотидів — триплет, чи кодон), неперекривність і наявність «стартових» кодонів, з яких починається зчитування (трансляція).
Також можна відзначити вкрай схожі процеси метаболізму в клітинах всіх організмів. Так, гліколіз у всіх еукаріотів і у більшості прокаріотів проходить за 10 однакових кроків, в одній і тій же послідовності, з використанням одних і тих же десяти ферментів (при тому, що можливі тисячі різних, але термодинамічно еквівалентних шляхів гліколізу)[137]. У всіх вивчених видів основним переносником енергії в клітині є аденозинтрифосфат (АТФ), хоча цю роль могли б виконувати сотні інших молекул[138][139].
2-га хромосома людини

У всіх представників родини гомінідів по 24 хромосоми, за винятком людей, у яких лише 23 хромосоми. Людська 2-га хромосома, згідно з широко визнаною точкою зору, є результатом злиття двох хромосом предків[140][141].
Докази злиття засновані на наступних фактах:
- Хромосома людини відповідає двом хромосомам мавп. Найближчий родич людини, бонобо, має практично ідентичні тим, що знаходяться у 2-й хромосомі людини, послідовності ДНК, але вони розміщуються на двох окремих хромосомах. Те саме вірне для більш далеких родичів: горили і орангутана[142][143].
- На хромосомі людини наявні рудиментарні центромери. Звичайно хромосома має тільки одну центромеру, але на довгому плечі 2-ї хромосоми спостерігаються залишки другої[144].
- Крім того, на хромосомі людини наявні рудиментарні теломери. Звичайно теломери знаходяться тільки на кінцях хромосом, але послідовності нуклеотидів, характерні для теломер, спостерігаються також і в середині 2-ї хромосоми[145].
Таким чином 2-га хромосома являє собою переконливий доказ еволюційного походження людей і інших мавп від спільного предка.
Ендогенні ретровіруси
Ендогенні ретровіруси являють собою сліди древніх вірусних інфекцій в ДНК. Ретровіруси (такі як ВІЛ і Т-лімфотропний вірус людини, який спричиняє лейкоз і лімфому) вбудовують власний код в геном зараженого організму. Звичайно після цього клітина починає продукувати нові копії вірусу[146], але в цьому процесі можливі збої: копіювання вбудованої вірусної послідовності пригнічується клітиною власника, але послідовність залишається в структурі хромосоми. Якщо цей процес відбувся із стовбуровою статевою клітиною, тоді таку послідовність будуть наслідувати нащадки інфікованої особини. Вірогідність незалежного вбудування в геном однакових вірусів на однакові позиції є нехтовно малою[147]. Це означає, що генетичний код одного і того ж ретровірусу може бути присутнім у двох тварин на одній і тій же позиції в ДНК тільки в тому випадку, якщо ці тварини походять від спільного предка.
Близько 1 % людського геному займають ендогенні ретровіруси, загалом таких послідовностей в ДНК кожної людини близько 30 000[148]. Деякі із цих вірусів зустрічаються тільки у людини. Інші послідовності зустрічаються тільки у шимпанзе і у людини (там самим підтверджується походження людини і шимпанзе від одного предка). Також є послідовності, які зустрічаються у горил, шимпанзе і людини, у орангутанів, горил, шимпанзе і людини, і так далі.[149][150][151] Розподілення ендогенних ретровірусів точно відповідає філогенетичному дереву.
Також можна навести приклад із родини котячих. У малих кішок (точніше — у таких тварин як очеретяний кіт, європейська лісова кішка, степова кішка і свійська кішка) знайдена спільна ретровірусна вставка. Ні у яких інших хижих цей ретроген не виявлений[150][152][153].
Біогеографічні докази
 | |
 | |
| Кактус astrophytum asterias із Техасу (зверху) і молочай Euphorbia obesa із Південної Африки (внизу). Два види живуть у схожих природних умовах і набули схожих форм за рахунок конвергентної еволюції. При цьому вони відносяться не тільки до різних родин, але й до різних рядів. Незважаючи на сприятливі умови, кактуси практично повністю відсутні в Африці. | |
Біогеографія — наука, яка вивчає закономірності географічного поширення тварин і рослин, а також характер фауни і флори окремих територій.
Якщо два види порівняно нещодавно виникли від однієї популяції, то вони, як правило, мешкають недалеко від ареалу цієї початкової популяції, що означає і недалеко одна від одної. Таким чином, географічний розподіл видів має бути сумісний з філогенетичним деревом. Якщо не приймати до уваги теорію еволюції, то розумно припустити, що види живуть в найбільш придатних для них умовах. Але територія еволюції передбачає, що має бути багато сприятливих для виду місць, в яких представники виду тим не менш відсутні у зв'язку із наявністю географічних бар'єрів.[154]
Саме так справи і йдуть на практиці. Серед ссавців Австралії переважають сумчасті. Плацентарні ссавці представлені китоподібними, ластоногими і рукокрилими (які могли перебратися до Австралії порівняно легко), а також гризунами, які з'являються в палеонтологічному літописі в міоцені, коли Австралія наблизилася до Нової Гвінеї. При цьому природні умови Австралії сприятливі і для інших видів ссавців. Наприклад, завезені на континент кролі європейські («дикі кролики») швидко розмножилися, широко розселилися і продовжують витісняти аборигенні види. В Австралії та на Новій Гвінеї, на півдні Південної Америки і в Африці зустрічаються нелітаючі безкілеві птахи, свистуни (зубасті жаби) і дводишні, в інших частинах світу вони відсутні. Середовище в пустелях Африки, Америки та Австралії дуже схоже, і рослини з однієї пустелі добре ростуть в іншій. Тим не менш, кактуси були виявлені тільки в Америці (за винятком Rhipsalis baccifera, вочевидь занесеного в Старий Світ перелітними птахами). Багато африканських і австралійських сукулентів (рослини, що мають спеціальні тканини для запасання води) зовні нагадують кактуси за рахунок конвергентної еволюції, але належать до інших рядів. Морські жителі східних та західних берегів Південної Америки різні, за винятком деяких молюсків, ракоподібних і голкошкірих, але на протилежних берегах Панамського перешийка живе близько 30 % одних і тих же видів риб, що пояснюється недавнім виникненням перешийка (близько 3 млн років тому)[155]. На більшості океанічних островів (острови, які ніколи не були з'єднані з материком) відсутні наземні ссавці, земноводні та інші тварини, які не здатні долати значні водні перешкоди. Видовий склад фауни таких островів бідний і є результатом випадкового занесення деяких видів, як правило птахів, рептилій, комах[156].
Географічний розподіл видів у минулому, який можна частково відновити по викопних рештках, також повинен відповідати філогенетичному дереву. Теорія дрейфу материків і теорія еволюції дозволяють передбачити, де слід шукати ті чи інші викопні рештки. Перші скам'янілості сумчастих знайдені в Північній Америці, їх вік становить близько 80 млн років. 40 млн років тому сумчасті вже були поширені в Південній Америці, але в Австралії, де вони зараз домінують, сумчасті з'явилися лише близько 30 млн років тому. Теорія еволюції передбачає, що австралійські сумчасті походять від американських. Відповідно до теорії дрейфу материків, 30-40 млн років тому Південна Америка та Австралія ще залишалися частиною Гондвани, великого континенту в південній півкулі, а між ними знаходилася майбутня Антарктида. На підставі двох теорій дослідники передбачили, що сумчасті мігрували з Південної Америки до Австралії через Антарктиду 30-40 млн років тому. Це передбачення справдилось: починаючи з 1982 року на острові Сеймур, розташованому недалеко від Антарктиди, було знайдено більше десяти викопних сумчастих віком 35-40 млн років[157][158].
Найближчі родичі сучасних людей — горили і шимпанзе — мешкають і Африці. Виходячи з цього, в 1872 році Чарлз Дарвін припустив, що і стародавніх предків людини слід шукати в Африці[159]. Чимало дослідників, таких як Луїс, Мері і Річард Лікі, Раймонд Дарт і Роберт Брум, прислухались до поради Дарвіна, і починаючи з 1920-х років в Африці було знайдено безліч проміжних форм між людиною і людиноподібними мавпами[160]. Якби викопних австралопітеків виявили, наприклад, в Австралії, а не в Африці, то уявлення про еволюцію гомінід довелося б переглядати[6][161][162][163].
В'юрки Дарвіна
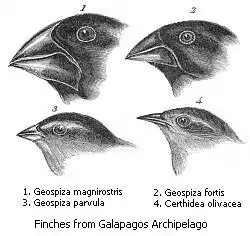
Під час навколосвітньої подорожі на кораблі «Бігль» Чарлз Дарвін описав 13 видів в'юрків, які мешкали на Галапагоських островах. Спостереження за цими птахами наштовхнуло Дарвіна на ідею походження видів за рахунок мінливості і природного відбору. Всі галапагоські в'юрки походять від спільного предка, який випадково потрапив сюди із Південної Америки. Від початкової форми, яка харчувалась насінням, виникли три групи птахів: земляні в'юрки, які харчуються насінням, комахоїдні деревні в'юрки і славкові в'юрки, які також харчуються дрібним насінням. Внаслідок пристосування до різних джерел їжі, в'юрки почали сильно відрізнятися один від одного будовою дзьоба. Три звичайних види в'юрків — великий, середній і малий — зустрічаються на більшості островів. В цьому випадку вони добре розрізняються за розмірами дзьобів і, відповідно, за розмірами насіння, якому вони надають перевагу. На одному із островів мешкає лише середній земляний в'юрок, і тут у птахів дзьоби менші — у відсутності конкурентів середній земляний в'юрок частково займає нішу малого в'юрка[164].
Див. також
Література
- Чарлз Дарвін. Походження видів. — 1859.
- Чарлз Дарвін. Походження людини і статевий відбір. — 1871.
- Верн Грант. Эволюционный процесс. — 1985. — ISBN 5-03-001432-2.
- Smith, J. M.. The Theory of Evolution. — Cambridge University Press, 1993. — ISBN 0521451280.
- Douglas J. Futuyma. Evolutionary Biology. — 3-е изд. — Sunderland, Massachusetts : Sinauer Associates, 1998. — ISBN 0-87893-189-9.
- Николай Иорданский. Эволюция жизни. — М. : Академия, 2001. — 432 с. — ISBN 5-7695-0537-0.
- Яблоков О. В., Юсуфов А. Г. Эволюционное учение: Учеб. для биол. спец. вузов. — М. : Высшая школа, 2006. — 310 с. — ISBN 5-06-004584-6.
- Jerry Coyne. Why Evolution is True. — Viking Adult, 2009. — ISBN 0670020532.
- Richrd Dawkins. The Greatest Show on Earth: The Evidence for Evolution. — 2009. — ISBN 0-593-06173-X.
- Происхождение жизни. Наука и вера CORPUS, Издательство «Астрель», 2010. ISBN 978-5-271-24664-7
Посилання
- Докази еволюції. Державний Дарвінівський музей. Архів оригіналу за 21 серпня 2011. Процитовано 7 вересня 2011.
- Джеймс Трефіл. Теорія еволюції. 200 законів світобудови. Архів оригіналу за 13 вересня 2011. Процитовано 7 вересня 2011.
- Бородін П. М. Еволюція — шляхи і механізми. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
- Докази еволюції. Загальна біологія: Навчальний посібник для 11-го класу. 2002. Архів оригіналу за 23 червня 2012. Процитовано 7 вересня 2011.
- Марков О. В. (ред.) (2010). Докази еволюції. Архів оригіналу за 5 вересня 2011. Процитовано 7 вересня 2011.
- Кондрашов А. С. (2010). Курс лекцій «Введення в еволюційну біологію». Архів оригіналу за 13 вересня 2013. Процитовано 7 вересня 2011.
Примітки
- Карл Воуз висунув гіпотезу, згідно з якою на перших етапах розвитку життя, коли формувалися перші клітини, основну роль відігравав горизонтальний перенос генів. В цьому випадку можливо, що останній універсальний спільний предок являв собою не одну клітину, а групу примітивних клітин, які вільно обмінювались генетичним матеріалом. (Карл Воуз. The universal ancestor // Proceedings of the National Academy of Sciences. — . — Т. 95, вип. 12. — С. 6854—6859.)
- Доказами еволюції тут вважаються не математично строгі докази (які можливі тільки в математиці), а свідчення, які дозволяють переконатися в істинності засад еволюційної теорії згідно із науковими методами
- Як і будь-яка інша модель, філогенетичне дерево відповідає дійсності не абсолютно точно. Горизонтальна передача генів і гібридизація ускладнюють використання дерев для опису зв'язків між видами. У тих рідкісних випадках, коли необхідно враховувати ці фактори, доводиться використовувати складніші моделі.
- Походження від спільного предка — генетичний процес, в якому «майбутнє» не залежить від «минулого» при відомому «теперішньому»: еволюція нащадків певної фіксованої популяції не залежить від того, які предки були у цієї початкової популяції. А значить, поступове еволюційне походження від спільного предка має відповідати таким моделям як марківські процеси і ланцюги Маркова. Використовуючи ці моделі, можна строго довести, що розгалужені марковскі системи утворюють багаторівневі ієрархічні структури. Тому біологи систематично використовують ланцюги Маркова для моделювання еволюційних процесів, в числі яких — складні генетичні процеси, зміна частотного розподілу прізвищ з часом і повединка патогенних мікроорганізмів при епідеміях.
- Перехідні форми між двома групами організмів не обов'язково мають бути нащадками однієї групи і предками іншої. По викопним решткам, як правило, неможливо достеменно встановити чи є один організм прямим предком іншого. Крім того, вірогідність знайти в палеонтологічному літописі чийого-небудь предка є дуже малою, і за замовчуванням серед біологів прийнято таку можливість взагалі не розглядати. Набагато більша вірогідність виявити відносно близьких родичів цього предка, схожих на нього будовою. Тому будь-яка перехідна форма автоматично інтерпретується як бічна гілка еволюції, а не «ділянка стовбура»Олександр Марков. Викопні риби в черговий раз підтвердили правоту Дарвіна. Элементы.ру. Архів оригіналу за 13 вересня 2011. Процитовано 7 вересня 2011.(рос.))
- Див. ілюстрацію до розділу Біогеографічні докази.
- Не слід плутати з «біогенетичним законом» Ернста Геккеля. Сумно відома ілюстрація із праці Геккеля, на якій, згідно із розповсюдженою точкою зору, відмінності між ембріонами штучно занижені. Карл Бер не був еволюціоністом, не використовував рисунки Геккеля, відкрив свій закон задовго до Геккеля і за 30 років до публікації «Походження видів». Сьогодні ембріологічні докази еволюції визнані науковою спільною, а «біогенетичний закон» — відкинутий.
- Не слід плутати з морськими свинками.
- Крім 20 стандартних амінокислот, в рідких випадках використовуються селеноцистеїн і пірролізин.
Джерела
- AAAS Council. (26 грудня 1922). AAAS Resolution: Present Scientific Status of the Theory of Evolution (англійською). Американська асоціація сприяння розвитку науки. Архів оригіналу за 27 вересня 2011. Процитовано 7 вересня 2011.
IAP Statement on the Teaching of Evolution (PDF) (англійською). Міжакадемічна група по міжнародним проблемам (IAP). 2006. Архів оригіналу за 29 вересня 2011. Процитовано 25 квітня 2007.
Рада директорів Американської асоціації сприяння розвитку науки. (16 лютого 2006). Statement on the Teaching of Evolution (PDF) (англійською). Американська асоціація сприяння розвитку науки. Архів оригіналу за 27 вересня 2011. Процитовано 7 вересня 2011.
Statements from Scientific and Scholarly Organizations (англійською). National Center for Science Education. Архів оригіналу за 15 серпня 2009. Процитовано 7 вересня 2011. - Observed Instances of Speciation (англійською). Архів TalkOrigins.org. Архів оригіналу за 2 вересня 2011. Процитовано 7 вересня 2011.
- Біологам вдалося побачити процес еволюції у дії. Lenta.ru. 11 липня 2008. Архів оригіналу за 5 липня 2011. Процитовано 1 грудня 2009.
- Морфологічні докази. Докази еволюції. Державний Дарвінівський музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Ембріологічні докази. Докази еволюції. Державний Дарвінівський музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Біогеографічні докази. Докази еволюції. Державний Дарвінівський музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Futuyma, 1998, p. 267—271, 283—294 (англ.)
- Genetic change (англійською). Архів TalkOrigins.org. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
- John Endler. Natural Selection in the Wild. — Принстон, Нью-Джерсі : Princeton University Press, 1986. — 336 с. — ISBN 0-691-00057-3. (англ.)
- Morphological change (англійською). Архів TalkOrigins.org. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
Barsh, G. S. The genetics of pigmentation: from fancy genes to complex traits // Trends in Genetics. — 1996. — Vol. 12, no. 8. — P. 299—305. DOI:10.1016/0168-9525(96)10031-7
Houde, A. E. (1988) «Genetic differences in female choice between two guppy populations.» Animal Behavior 36: 510—516. (англ.)
Morton, E. S. (1990) «Habitat segregation by sex in the hooded warbler: Experiments on proximate causation and discussion of its evolution.» American Naturalist 135: 319—333. (англ.)
Johnston, R. F., and Selander, R. K. (1973) «Evolution in the house sparrow. III. Variation in size and sexual dimorphism in Europe and North and South America.» American Naturalist 107: 373—390. (англ.)
Futuyma, 1998, p. 247—262 (англ.) - Functional change (англійською). Архів TalkOrigins.org. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
-
Luria, S., and Delbruck, M. (1943) «Mutations of bacteria from virus sensitivity to virus resistance.» Genetics 28: 491—511. (англ.)
Lederberg, J., and Lederberg, E. M. (1952) «Replica plating and indirect selection of bacterial mutants.» Journal of Bacteriology 63: 399—406. (англ.)
Bennett, A. F., R. E. Lenski, et al. Evolutionary adaptation to temperature. I. Fitness responses of Escherichia coli to changes in its thermal environment // Evolution. — 1992. — Vol. 46. — P. 16—30.
Ffrench-Constant, R. H., N. Anthony, et al. Cyclodiene insecticide resistance: from molecular to population genetics // Annual Review of Entymology. — 2000. — Vol. 45. — P. 449—466. DOI:10.1146/annurev.ento.45.1.449
Carpenter, M. A. and O’Brien, S. J. Coadaptation and immunodeficiency virus: lessons from the Felidae // Current Opinion in Genetics and Development. — 1995. — Т. 5. — P. 739—745.
Richter, T. E., and Ronald, P. C. The evolution of disease resistance genes // Plant Molecular Biology. — 2000. — Vol. 42. — P. 195—204. DOI:10.1023/A:1006388223475
Andren, C., M. Marden, et al. (1989) «Tolerance to low pH in a population of moor frogs, Rana arvalis from an acid and a neutral environment: a possible test case of rapid evolutionary response to acidification.» Oikos 56: 215—223. (англ.)
Bishop, J. A. (1981) «A neo-Darwinian approach to resistance: Examples from mammals.» In Genetic Consequences of Man Made Change. J. A. Bishop and L. M. Cook. London, Academic Press. (англ.) - Okada, H., S. Negoro, et al. Evolutionary adaptation of plasmid-encoded enzymes for degrading nylon oligomers // Nature. — 1983. — Vol. 306, no. 5939. — P. 203—206. DOI:10.1038/306203a0
Orser, C. S., and Lange, C. C. Molecular analysis of pentachlorophenol degradation // Biodegradation. — 1994. — Vol. 5. — P. 277—288.
Lee, S. G., B. D. Yoon, et al. Isolation of a novel pentachlorophenol-degrading bacterium, Pseudomonas sp. Bu34 // Journal of Applied Microbiology. — 1998. — Vol. 85. — P. 1—8.
S Negoro, S Kakudo, I Urabe, and H Okada. A new nylon oligomer degradation gene (nylC) on plasmid pOAD2 from a Flavobacterium sp // J Bacteriol. — 1992. — Vol. 174, № 24. — P. 7948—7953. - Bob Holmes. (9 июня 2008). Bacteria make major evolutionary shift in the lab (англійською). New Scientist. Архів оригіналу за 9 жовтня 2008. Процитовано 7 вересня 2011.
- Шапошников Г.Х. Специфичность и возникновение адаптации к новым хозяевам у тлей (Homoptera, Aphidoidea) в процессе естественного отбора (экспериментальное исследование) // Энтом. обозр.. — 1961. — Т. XL, вип. 4. — С. 739—762.
- Шапошников Г.Х. Морфологическая дивергенция и конвергенция в эксперименте с тлями (Homoptera, Aphidinea) // Энтом. обозр.. — Т. XLIV, № 1. — С. 3—25.
- Шапошников Г.Х. Возникновение и утрата репродуктивной изоляции и критерий вида // Энтом. обозр.. — 1966. — Т. XLV, № 1. — С. 3—35.
- Шапошников Г.Х. Динамика клонов, популяций и видов и эволюция // Журн. общ. биологии. — 1978. — Т. XXXIX, вип. 1. — С. 15—33.
- Neil S. Greenspan. HIV Drug Resistance: Problems and Perspectives // Infect. Dis. Rep.. — 2013. — Т. 5. — С. e5. — DOI:.
- UMass Amherst : In the Loop : Talking Points : Lizards undergo rapid evolution after introduction to new home, says researcher (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 27 травня 2008.
- PZ Myers. Pharyngula: Still just a lizard (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 27 травня 2008.
- Herrel A, Huyghe K, Vanhooydonck B, et al. Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource // Proc. Natl. Acad. Sci. USA. — 2008. — Т. 105, № 12 (березень). — С. 4792—5. — PMID:. DOI:10.1073/pnas.0711998105
Ящірки стрімко еволюціонують в нових умовах. globalscience.ru. Архів оригіналу за 23 червня 2011. Процитовано 7 вересня 2011.
National Geographic: Lizards Rapidly Evolve After Introduction to Island (англійською). Архів оригіналу за 11 травня 2008. Процитовано 27 травня 2008.
Science Daily: Lizards Undergo Rapid Evolution After Introduction To A New Home (англійською). Архів оригіналу за 17 травня 2008. Процитовано 27 травня 2008.
Newswise: Lizards Undergo Rapid Evolution After Introduction to New Island (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 6 серпня 2008.
Bart Vervust, Irena Grbac, Raoul Van Damme. (August 2007). Differences in morphology, performance and behaviour between recently diverged populations of Podarcis sicula mirror differences in predation pressure. Oikos (англійською) 116 (8): 1343—1352. doi:10.1111/j.0030-1299.2007.15989.x.
Dawkins, 2009, p. 113—116 (англ.) - Lizards Rapidly Evolve After Introduction to Island. news.nationalgeographic.com. Процитовано 8 січня 2017.
- Tanaka T, Hashimoto H. (1989). Drug-resistance and its transferability of Shigella strains isolated in 1986 in Japan. Kansenshogaku Zasshi 63 (1): 15—26. (англ.)
- Олександр Марков. (2007). Комахи-шкідники захищаються від біологічної зброї. Архів оригіналу за 18 жовтня 2011. Процитовано 7 вересня 2011.
S. Asser-Kaiser, E. Fritsch, et al. Rapid Emergence of Baculovirus Resistance in Codling Moth Due to Dominant, Sex-Linked Inheritance // Science. — 2007. — Т. 317, № 5846. DOI:10.1126/science.1146542 - Вид (в биологии) // Велика радянська енциклопедія.
- Speciation stages (англійською). Архів TalkOrigins.org. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
- Futuyma, 1998, p. 454 (англ.)
- Irwin, D. E., Irwin, J. H., and Price, T. D. Ring species as bridges between microevolution and speciation. — 2001. — Vol. 112—113. — P. 223—243. DOI:10.1023/A:1013319217703
-
Grant, 1985, Перекрывающиеся кольца рас. Архівовано 27 вересня 2009 у Wayback Machine.
Moritz, C., C. J. Schneider, et al. (1982) «Evolutionary relationships within the Ensatina eschscholtzii complex confirm the ring species interpretation.» Systematic Biology 41: 273—291. (англ.)
Futuyma, 1998, p. 455—456 (англ.) - Barton, N. H., and Gale, R. S. (1993) «Genetic analysis of hybrid zones.» In Hybrid Zones and the Evolutionary Process. New York, Oxford University Press: 12—45.
- Smith, 1993, p. 253, 264 (англ.)
- Futuyma, 1998, p. 460 (англ.)
- Smith, 1993; Futuyma, 1998, главы 15 и 16 (англ.)
- Adaptive Traits of the Polar Bear (Ursus Maritimus) (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Polar Bear Evolution (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Ron Rayborne Accepts Hovind’s Challenge (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Futuyma, 1998, p. 510 (англ.)
- Gingerich, P. D.. // Science. — Vol. 222, no. 4620. — P. 159—161. DOI:10.1126/science.222.4620.159
- Speciations (англійською). Архів TalkOrigins.org. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
- Pasterniani, E. Selection for reproductive isolation between two populations of maize, Zea mays L // Evolution. — 1969. — Т. 23. — С. 534—547.
- Gottlieb, L. D. Genetic differentiation, sympatric speciation, and the origin of a diploid species of Stephanomeria // American Journal of Botany. — 1973. — Т. 60. — С. 545—553.
- Boraas, M. E. 1983. Predator induced evolution in chemostat culture. EOS. Transactions of the American Geophysical Union. 64:1102. (англ.)
- Shikano, S., L. S. Luckinbill, Y. Kurihara. // Microbial Ecology. — 1990. — Vol. 20, no. 1. — P. 75—84. DOI:10.1007/BF02543868
- Britton-Davidian, J., J. Catalan, et al. Rapid chromosomal evolution in island mice // Nature. — 2000. — Vol. 403. — P. 158. DOI:10.1038/35003116
- As the Worm Turns: Speciation and the Apple Maggot Fly (англійською). National Center for Case Study Teaching in Science. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Feder JL, Roethele JB, Filchak K, Niedbalski J, Romero-Severson J. (1 березня 2003). Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella. Genetics 163 (3): 939—53. PMC 1462491. PMID 12663534. (англ.)
- Цепная реакция видообразования. Элементы.ру. Архів оригіналу за 7 жовтня 2011. Процитовано 7 вересня 2011.
- Для видоутворення достатньо одного гена. Элементы.ру. Архів оригіналу за 7 жовтня 2011. Процитовано 7 вересня 2011.
- Grant, 1985, Смежно-симпатрическое видообразование. Архівовано 13 вересня 2009 у Wayback Machine.
-
Berlocher SH, Bush GL. (1982). An electrophoretic analysis of Rhagoletis (Diptera: Tephritidae) phylogeny. Systematic Zoology 31: 136—55. doi:10.2307/2413033. (англ.)
Berlocher SH, Feder JL. (2002). Sympatric speciation in phytophagous insects: moving beyond controversy?. Annu Rev Entomol. 47: 773—815. PMID 11729091. doi:10.1146/annurev.ento.47.091201.145312. Архів оригіналу за 31 липня 2010. Процитовано 12 квітня 2019. (англ.)
Bush GL. (1969). Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera: Tephritidae). Evolution 23: 237—51. doi:10.2307/2406788. (англ.)
Prokopy RJ, Diehl SR, Cooley SS. (June 1988). Behavioral evidence for host races in Rhagoletis pomonella flies. Oecologia 76 (1): 138—47. doi:10.1007/BF00379612. (англ.)
Feder JL, Roethele JB, Wlazlo B, Berlocher SH. (October 1997). Selective maintenance of allozyme differences among sympatric host races of the apple maggot fly. Proc Natl Acad Sci USA. 94 (21): 11417—21. PMC 23485. PMID 11038585. doi:10.1073/pnas.94.21.11417. (англ.) - Дарвін як приклад використовував групування зірок в сузір'я: Дарвин, 1859. Глава XIV. Классификация.
- Swofford, D. L., Olsen, G. J., Waddell, P. J., and Hillis, D. M. (1996) «Phylogenetic inference.» In Molecular Systematics, pp 407—514. Hillis, D. M., Moritz, C. and Mable, B. K. eds., Sunderland, Massachusetts: Sinauer, p. 504 (англ.)
-
Archie, J. W. (1989) «A randomization test for phylogenetic information in systematic data.» Systematic Zoology 38: 219—252. (англ.)
Faith, D. P., and Cranston, P. S. (1991) «Could a cladogram this short have arisen by chance alone?: on permutation tests for cladistic structure.» Cladistics 7: 1—28. (англ.)
Farris, J. S. (1989) «The retention index and the rescaled consistency index.» Cladistics 5:417—419. (англ.)
Felsenstein, J. (1985) «Confidence limits on phylogenies: an approach using the bootstrap.» Evolution 39: 783—791. (англ.)
Huelsenbeck, J. P., Ronquist, F., Nielsen, R., and Bollback, J. P. Bayesian inference of phylogeny and its impact on evolutionary biology // Science. — 2001. — Vol. 294, no. 5550. — P. 2310—2314. DOI:10.1126/science.1065889 -
Hillis, D. M. (1991) «Discriminating between phylogenetic signal and random noise in DNA sequences.» In Phylogenetic analysis of DNA sequences. pp. 278—294 M. M. Miyamoto and J. Cracraft, eds. New York: Oxford University Press. (англ.)
Hillis, D. M., Huelsenbeck, J. P. Signal, noise, and reliability in molecular phylogenetic analyses // Journal of Heredity. — 1992. — Vol. 83. — P. 189—195.
Klassen, G. J., Mooi, R. D., and Locke, A. (1991) «Consistency indices and random data.» Syst. Zool. 40:446—457. (англ.) -
Baldauf, S. L., Roger, A. J., Wenk-Siefert, I., and Doolittle, W. F. A kingdom-level phylogeny of eukaryotes based on combined protein data // Science. — Vol. 290, no. 5493. — P. 972—977. DOI:10.1126/science.290.5493.972
Brown, J. R., Douady, C. J., Italia, M. J., Marshall, W. E., Stanhope, M. J. Universal trees based on large combined protein sequence data sets // Nature Genetics. — 2001. — Vol. 28. — P. 281—285. DOI:10.1038/90129 - Nested hierarchy (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Penny, D., Foulds, L. R., and Hendy, M. D. (1982) «Testing the theory of evolution by comparing phylogenetic trees constructed from five different protein sequences.» Nature 297: 197—200.
Penny, D., Hendy, M. D., and Steel, M. A. (1991) «Testing the theory of descent.» In Phylogenetic Analysis of DNA Sequences, eds. Miyamoto, M. and Cracraft, J., New York: Oxford University Press. pp. 155—183.
Zuckerkandl, E. and Pauling, L. (1965) «Evolutionary Divergence and Convergence in Proteins.» in Evolving Genes and Proteins: a symposium held at the Institute of Microbiology of Rutgers, with support from the National Science Foundation. Eds Vernon Bryson and Henry J. Vogel. New York: Academic Press. (англ.) - Independent convergence (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Fossil steroids record the appearance of Demospongiae during the Cryogenian period // Nature. — Vol. 457. — P. 718—721. DOI:10.1038/nature07673
- Самые древние позвоночные животные. Державний Дарвінівський музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Earth’s strange past and the fossil record (англійською). Архів TalkOrigins.org. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
- Morphological intermediates (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Hu, D.; Hou, L.; Zhang, L. & Xu, X. (2009). A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461 (7264): 640—643. PMID 19794491. doi:10.1038/nature08322. Вказано більш, ніж один
|last1=та|last=(довідка); Вказано більш, ніж один|first1=та|first=(довідка) (англ.) - Див. неповний список перехідних форм в статті Список перехідних форм.
- Dawkins, 2009, p. 151 (англ.)
- Dawkins, 2009, p. 149 (англ.)
- Річард Докінз. Какая жалость — честный креационист!. Архів оригіналу за 14 жовтня 2011. Процитовано 7 вересня 2011.
- The La Brea Exploration Guide (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Pamela J. W. Gore. Laboratory 11 — Fossil Preservation (англійською). Georgia Perimeter College. Архів оригіналу за 28 серпня 2008. Процитовано 7 вересня 2011.
- Палеонтологические доказательства. Докази еволюції. Державний Дарвінівський музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Dawkins, 2009, p. 187 (англ.)
- Дональд Джохансон, Мейтленд Иди. Глава 12 // Люси: истоки рода человеческого. — М., 1984.
- Dawkins, 2009, p. 189 (англ.)
- Dawkins, 2009, p. 188 (англ.)
- Древнейшим предком человека признали ардипитека Арди. Lenta.ru. Архів оригіналу за 31 березня 2012. Процитовано 7 вересня 2011.
Jamie Shreeve. (1 жовтня 2009). Oldest Skeleton of Human Ancestor Found: Move over, Lucy. And kiss the missing link goodbye. National Geographic. Архів оригіналу за 4 жовтня 2009. Процитовано 1 жовтня 2009.
Ann Gibbons. Ancient Skeleton May Rewrite Earliest Chapter of Human Evolution (англійською). Science. Архів оригіналу за 4 жовтня 2009. Процитовано 1 жовтня 2009. (англ.) - White, Tim D.; Asfaw, Berhane; Beyene, Yonas; Haile-Selassie, Yohannes; Lovejoy, C. Owen; Suwa, Gen; WoldeGabriel, Giday (2009). Ardipithecus ramidus and the Paleobiology of Early Hominids. Science 326 (5949): 75—86. doi:10.1126/science.1175802. (англ.)
- Fossil finds extend human story. BBC. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011. (англ.)
- Див. Список викопних рештків людської еволюції
- Походження і еволюція людини. evolbiol.ru. Архів оригіналу за 15 жовтня 2011. Процитовано 7 вересня 2011.
- Чарльз Дарвин. Глава VII // Происхождение человека и половой отбор.
- Dawkins, 2009, p. 194 (англ.)
- Еволюція кінцівок коня. Державний Дарвінівський музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Эволюционная палеонтология. Становление эволюционной биологии. Державний Дарвінівський музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Воронцов Н. Н., Сухорукова Л. Н. Эволюция органического мира. Учебное пособие для 10—11 кл. ср. шк. — 2-е изд., перераб. и доп. — М. : Наука. — 256 с. — 15 000 прим. — ISBN 5020060437. (рос.)
- Morphological analogy (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Morphological parahomology (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Дарвин, 1859. Глава 14. Морфология.
- Dawkins, 2009, p. 297 (англ.)
- Crapo, R. (1985) «Are the vanishing teeth of fetal baleen whales useless?» Origins Research 7: 1. (англ.)
Culver, D. C., Fong, D. W., and Kane T. C. (1995) «Vestigialization and Loss of Nonfunctional Characters.» Annual Review of Ecology and Systematics 26: 249—268. (англ.)
Дарвин, 1859. Глава XIV. Рудиментарные, атрофированные и абортивные органы. «Орган, який слугує для двох цілей, може стати рудиментарним і навіть абсолютно недорозвиненим для однієї, навіть важливішої мети і залишитися цілком придатним для іншої», «орган може стати рудиментарним для своєї спеціальної функції і вживатися для зовсім іншої мети».
Dodson, E. O. (1960) Evolution: Process and Product. New York: Reinhold Publishers, стр. 44. (англ.)
Griffiths, P. (1992) «Adaptive explanation and the concept of a vestige.» in Trees of Life: essays in philosophy of biology. Edited by Paul Griffiths. Dordrecht, Boston: Kluwer Academic Publishers. (англ.)
Hall, B. K. Descent with modification: the unity underlying homology and homoplasy as seen through an analysis of development and evolution // Biol Rev Camb Philos Soc. — 2003. — Vol. 78, no. 3. — P. 409—433. DOI:10.1017/S1464793102006097
McCabe, J. (1912) The Story of Evolution. Boston: Small, Maynard, and Co, стр. 264. (англ.)
Merrell, D. J. (1962) Evolution and Genetics: The Modern Theory of Evolution. New York: Holt, Rinehart and Winston, стр. 101. (англ.)
Moody, P. A. (1962) Introduction to Evolution. Second edition. New York: Harper, стр. 40. (англ.)
Muller, G. B. (2002) «Vestigial Organs and Structures.» in Encyclopedia of Evolution. Mark Pagel, editor in chief, New York: Oxford University Press. (англ.)
Naylor, B. G. (1982) «Vestigial organs are evidence of evolution.» Evolutionary Theory 6: 91—96. (англ.)
Strickberger (2000) Evolution. Third edition. Sudbury, MA: Jones and Bartlett. (англ.)
Weismann, A. (1886) «IX. Retrogressive Development in Nature.» reproduced in Essays upon Heredity and Kindred Biological Problems. Volume II. Poulton, E. B. and Shipley, A. E., editors., Clarendon Press: Oxford. 1892. Стр. 9—10. (англ.)
Wiedersheim, R. (1893) The Structure of Man: An Index to His Past History. Second Edition. Translated by H. and M. Bernard. London: Macmillan and Co. 1895. Стр. 2, 200, 205. (англ.) - Дарвин, 1859. Глава XIV. Рудиментарные, атрофированные и абортивные органы. «Неможливо назвати будь-яку з вищих тварин, у якої та чи інша частина не була б у рудиментарному стані.»
- Дарвин, 1859. Глава XIV. Рудиментарные, атрофированные и абортивные органы. «Крило пінгвіна має велике значення, діючи як плавник, отже, воно може являти собою і ранню стадію утворення крила, хоча я цього не думаю, більше ймовірно, що це редукований орган, модифікований для нової функції, з іншого боку, крило ківі зовсім не має користі і дійсно рудиментарне.»
- Жан Батист Ламарк. Філософія зоології. — 1809. «Сліпак Олів'є, який веде подібно кроту, підземний спосіб життя і, ймовірно, ще рідше, ніж кріт, що піддається дії денного світла, зовсім перестав користуватися зором, і в нього збереглися лише сліди цього органу, але навіть ці сліди зовсім сховані під шкірою і деякими іншими покриваючими їх частинами, які не залишають жодного доступу для світла.»
- Рудиментарні органи // Велика радянська енциклопедія.
- Grant, 1985, Докази еволюції. Архівовано 10 листопада 2009 у Wayback Machine.
- Cohn, M. J., and Tickle, C. Developmental basis of limblessness and axial patterning in snakes // Nature. — 1999. — Vol. 399. — P. 474—479. Cohn, M. J. Developmental mechanisms of vertebrate limb evolution // Novartis Found Symp. — 2001. — Vol. 232. — P. 47—57.
- Дарвин, 1859. Глава XIV. Рудиментарные, атрофированные и абортивные органы.
- Vestiges (англійською). Архів TalkOrigins.org. Архів оригіналу за 29 жовтня 2006. Процитовано 7 вересня 2011.
- Дельфин встал и пошел. Итоги. Архів оригіналу за 5 березня 2016. Процитовано 7 вересня 2011.
- http://universe-review.ca/I10-10-snake.jpg
- Tyson, Reid; Graham, John P.; Colahan, Patrick T.; Berry, Clifford R. (июль 2004). Skeletal Atavism in a Miniature Horse. Veterinary Radiology & Ultrasound 45 (4): 315—317. (англ.)
- Клещи вновь открыли радость секса. Инновационный портал Уральского Федерального округа. Архів оригіналу за 13 березня 2016. Процитовано 7 вересня 2011.
- Domes, Katja; Norton, Roy A.; Maraun, Mark & Scheu, Stefan. Reevolution of sexuality breaks Dollo's law // PNAS. — 2007. — Vol. 104, iss. 17. — P. 7139—7144.
- Atavisms (англійською). Архів TalkOrigins.org. Архів оригіналу за 29 жовтня 2006. Процитовано 7 вересня 2011.
- П. М. Бородин. Возникновение адаптаций в результате естественного отбора. Эволюция — пути и механизмы. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Н. М. Борисов, Ф. Ю. Воробьев, А. М. Гиляров, К. Ю. Еськов, А. Ю. Журавлев, А. В. Марков, А. А. Оскольский, П. Н. Петров, А. Б. Шипунов. (2010). Доказательства эволюции. Архів оригіналу за 5 вересня 2011. Процитовано 7 вересня 2011.
Mark Ridley. Evolution. — 3-е изд. — Blackwell Publishing, 2004. — P. 282. — ISBN 1405103450. (англ.)
The Giraffe: Inside Nature’s Giants (англійською).
Dawkins, 2009, p. 361 (англ.) - Goldsmith, T. H. Optimization, constraint, and history in the evolution of eyes // Quarterly Review of Biology. — 1990. — Vol. 65. — P. 281—322.
Williams, G. C. (1992) Natural Selection: Domains, Levels, and Challenges. New York, Oxford University Press, pp. 72—74. (англ.) - Morphological inefficiency (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Evidence for Jury-Rigged Design in Nature (англійською). Архів TalkOrigins.org. Архів оригіналу за 4 вересня 2011. Процитовано 7 вересня 2011.
- Річард Докінз. Поклоніння «білим плямам» // Бог як ілюзія.
- Станіслав Сковрон. Порівняльно-ембріологічні докази еволюції. Розвиток теорії еволюції. Архів оригіналу за 9 січня 2011. Процитовано 7 вересня 2011.
- Иорданский, 2001, Глава 3. Мезозойська ера — час рептилій. Архівовано 25 травня 2009 у Wayback Machine.
- Gilbert, S. F. (1997) Developmental Biology. Fifth edition. Sinauer Associates, pp. 380, 382 (англ.)
- Kardong, K. V. (2002) Vertebrates: Comparative Anatomy, Function, Evolution. Third ed. New York: McGraw Hill, pp. 52, 504, 581. (англ.)
- Raynaud, A. Developmental mechanism involved in the embryonic reduction of limbs in reptiles // Int J Dev Biol. — 1990. — Vol. 34. — P. 233—243.
Raynaud, A., Kan, P. DNA synthesis decline involved in the developmental arrest of the limb buds in the embryos of the slow worm, Anguis fragilis (L.) // Int J Dev Biol. — 1992. — Vol. 36. — P. 303—310.
Raynaud, A., Van den Elzen, P. Rudimentary stages of the extremities of Scelotes gronovii (Daudin) embryos, a South African Scincidea reptile. — 1976. — Vol. 65. — P. 17—36. - Amasaki, H., Ishikawa, H., Daigo, M. Developmental changes of the fore- and hind-limbs in the fetuses of the southern minke whale, Balaenoptera acutorostrata // Anat Anz. — 1989. — Vol. 169. — P. 145—148.
Sedmera, D., Misek, I., Klima, M. On the development of Cetacean extremities: I. Hind limb rudimentation in the Spotted dolphin Stenella attenuata // Eur J Morphol. — 1997. — Vol. 35. — P. 25—30. - Fallon, J. F., Simandl, B. K. Evidence of a role for cell death in the disappearance of the embryonic human tail. — 1978. — Vol. 152. — P. 111—129.
- Moore, K. L. and Persaud, T. V. N. (1998) The developing human: clinically oriented embryology. 6th ed., editor Schmitt, W., Saunders, Philadelphia, pp. 91—100. (англ.)
- Nievelstein, R. A., Hartwig, N. G., Vermeij-Keers, C., Valk, J. Embryonic development of the mammalian caudal neural tube. — 1993. — Vol. 48. — P. 21—31.
- Sapunar, D., Vilovic, K., England, M., Saraga-Babic, M. Morphological diversity of dying cells during regression of the human tail // Ann Anat. — 2001. — Vol. 183. — P. 217—222.
- Saraga-Babic, M., Lehtonen, E., Svajger, A., Wartiovaara, J. // Anat Anz. — Vol. 176. — P. 277—286.
Saraga-Babic, M., Stefanovic, V., Saraga, M., Wartiovaara, J., Lehtonen, E. Expression of intermediate filaments and desmosomal proteins during differentiation of the human spinal cord. — 2002. — Vol. 104. — P. 157—166. - Ontogeny (англійською). Архів TalkOrigins.org. Архів оригіналу за 29 жовтня 2006. Процитовано 7 вересня 2011.
- П. М. Бородін. Ембріологічні свідоцтва еволюції. Еволюція — шляхи і механізми. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Джеймс Трефіл. Теорія еволюції. 200 законов мироздания. Архів оригіналу за 13 вересня 2011. Процитовано 7 вересня 2011.
- Chimpanzee Sequencing and Analysis Consortium. (2005). Initial sequence of the chimpanzee genome and comparison with the human genome (PDF). Nature 437: 69—87. doi:10.1038/nature04072. (September 1). (англ.)
Cheng Z, Ventura M et al. (2005). A genome-wide comparison of recent chimpanzee and human segmental duplications. Nature 437: 88—93. doi:10.1038/nature04000. (September 1). (англ.) - Stewart, C. B., Disotell, T. R. Primate evolution—in and out of Africa // Current Biology. — 1998. — Vol. 8. — P. R582—R588. DOI:10.1016/S0960-9822(07)00367-3
- Crowe, J. F. (1993) «Mutation, fitness, and genetic load.» Oxford Survey of Evolutionary Biology 9: 3—42.
Futuyma, 1998, p. 273 (англ.) - Giannelli, F., Anagnostopoulos, T., Green, P. M. Mutation rates in humans. II. Sporadic mutation-specific rates and rate of detrimental human mutations inferred from hemophilia B // Am J Hum Genet. — 1999. — Vol. 65. — P. 1580—1587.
Mohrenweiser, H. Impact of the molecular spectrum of mutational lesions on estimates of germinal gene-mutation rates. — 1994. — Vol. 304. — P. 119—137. - Genetic rates of change (англійською). Архів TalkOrigins.org. Архів оригіналу за 25 вересня 2011. Процитовано 7 вересня 2011.
-
Rozenski, J., Crain, P. F. et al. The RNA Modification Database: 1999 update // Nucleic Acids Research. — 1999. — Vol. 27. — P. 196—197.
Voet, D., and Voet, J. (1995) Biochemistry. New York: John Wiley and Sons, p. 969 (англ.) - Voet, D., and Voet, J. (1995) Biochemistry. New York: John Wiley and Sons, p. 69. (англ.)
Garavelli, J. S., Hou, Z., et al. The RESID Database of protein structure modifications and the NRL-3D Sequence-Structure Database // Nucleic Acids Research. — 2001. — Vol. 29. — P. 199—201. - Yockey, H. P. (1992) Information Theory and Molecular Biology. New York: Cambridge University Press. (англ.)
- Ralph T. Hinegardner, Joseph Engelberg. Rationale for a Universal Genetic Code // Science. — 22 листопада 1963. — Vol. 142, no. 3595. — P. 1083—1085. DOI:10.1126/science.142.3595.1083
- Judson, H. F. (1996) The Eighth Day of Creation: Makers of the Revolution in Biology. Expanded edition. Cold Spring Harbor Press, p. 280—281. (англ.)
- Brenner, S. On the impossibility of all overlapping triplet codes in information transfer from nucleic acid to proteins // PNAS. — 1957. — Vol. 43, no. 8. — P. 687—694.
- Crick, F. H. C., Barnett, L., Brenner, S., and Watts-Tobin, R. J. (1961) «General nature of the genetic code for proteins.» Nature 192:1227—1232. (англ.)
- Voet, D., and Voet, J. (1995) Biochemistry. New York: John Wiley and Sons, p. 445. (англ.)
- The fundamental unity of life (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Біохімічні докази. Докази еволюції. Державний Дарвінівський музей музей. Архів оригіналу за 20 серпня 2011. Процитовано 7 вересня 2011.
- Alec MacAndrew. Human Chromosome 2 is a fusion of two ancestral chromosomes. Alec’s Evolution Pages (англійською). Архів оригіналу за 6 вересня 2011. Процитовано 4 серпня 2009.
- Evidence of Common Ancestry: Human Chromosome 2 (англійською). YouTube. Процитовано 4 серпня 2009.
- Yunis J. J., Prakash O. The origin of man: a chromosomal pictorial legacy // Science. — 19 марта 1982. — Vol. 215, № 4539. — P. 1525—1530. DOI:10.1126/science.7063861. PMID 7063861.
- Comparison of the Human and Great Ape Chromosomes as Evidence for Common Ancestry. The Evolution Evidence Page (англійською). Архів оригіналу за 20 серпня 2011. Процитовано 4 серпня 2009.
- Avarello R., Pedicini A., Caiulo A., Zuffardi O., Fraccaro M. Evidence for an ancestral alphoid domain on the long arm of human chromosome 2 // Human Genetics. — Springer Berlin/Heidelberg, май 1992. — Vol. 89, № 2. — P. 247—249. DOI:10.1007/BF00217134. PMID 1587535.
- IJdo J. W. Origin of human chromosome 2: an ancestral telomere-telomere fusion // Proceedings of the National Academy of Sciences. — National Academy of Sciences, 15 жовтня 1991. — Vol. 88, № 20. — P. 9051—9055. DOI:10.1073/pnas.88.20.9051. PMID 1924367.
- Анимация: жизненный цикл ретровируса (англійською). Архів оригіналу за 24 лютого 2014. Процитовано 7 вересня 2011.
- Barbulescu M, Turner G, Seaman MI, Deinard AS, Kidd KK, Lenz J. Many human endogenous retrovirus K (HERV-K) proviruses are unique to humans // Current Biology. — 1999. — Vol. 9, № 16. — P. 861—868.
- Е. Д. Свердлов. Retroviruses and primate evolution (англійською). Архів оригіналу за 5 січня 2013. Процитовано 7 вересня 2011.
- М. В. Супотницкий. ВИЧ/СПИД пандемия — так с чем же в действительности мы столкнулись?. Архів оригіналу за 5 жовтня 2011. Процитовано 7 вересня 2011.
- Endogenous retroviruses (англійською). Архів TalkOrigins.org. Архів оригіналу за 3 вересня 2011. Процитовано 7 вересня 2011.
- Lebedev et al. Differences in HERV-K LTR insertions in orthologous loci of humans and great apes // Gene. — 2000. — Vol. 247, no. 1—2. — P. 265—277. DOI:10.1016/j.physletb.2003.10.071
- Futuyma, 1998, p. 293—294 (англ.)
- Todaro, G. J., Benveniste, R. E., Callahan, R., Lieber, M. M., Sherr, C. J. Endogenous primate and feline type C viruses // Cold Spring Harb Symp Quant Biol. — 1975. — Vol. 39 Pt 2. — P. 1159—1168.
- Futuyma, 1998, p. 201—203 (англ.)
- Present biogeography (англійською). Архів TalkOrigins.org. Архів оригіналу за 29 жовтня 2006. Процитовано 7 вересня 2011.
- Острівна фауна // Велика радянська енциклопедія.
- Woodburne, M. O., and Case, J. A. (1996) «Dispersal, vicariance, and the Late Cretaceous to Early Tertiary land mammal biogeography from South America to Australia.» Journal of Mammalian Evolution 3: 121—161.
- Coyne, 2009, p. 91—99 (англ.)
- Чарлз Дарвін. Походження людини і статевий відбір.
- Кейт Вонг. Зов далекого предка // В мире науки. — 2003. — № 5. — С. 38—47.
- Дарвин, 1859. Глава 12. Географічне поширення.
Дарвин, 1859. Глава 13. Географічне поширення (продовження). - Географічне видоутворення. Державний Дарвінівський музей. Архів оригіналу за 21 серпня 2011. Процитовано 7 вересня 2011.
- Past biogeography (англійською). Архів TalkOrigins.org. Архів оригіналу за 29 жовтня 2006. Процитовано 7 вересня 2011.
- Географічне видоутворення. В'юрки Дарвіна. Державний Дарвінівський музей. Архів оригіналу за 21 серпня 2011. Процитовано 7 вересня 2011.

{kind=link}