Перехідна форма
Перехідні (проміжні) форми — організми, які поєднують в своїй будові ознаки двох великих систематичних груп.
| Частина серії статей на тему: |
| Еволюційна біологія |
|---|
 Лондонський екземпляр археоптерикса, першої знахідки перехідної форми між плазунами і птахами |
|
|
Процеси |
|
Розвиток теорії
|
|
Дисципліни і застосування
|
|
Вплив на соціум
|
|
|
Перехідні форми характеризуються наявністю більш стародавніх і примітивних (у значенні первинних) рис, ніж пізніші форми, але, у той же час, наявністю більш прогресивних (у значенні пізніших) рис, ніж їхні предки. Як правило, термін «перехідні форми» вживають по відношенню до викопних форм, хоча проміжні види зовсім не обов'язково повинні вмирати.
Перехідні форми використовують як один з доказів існування біологічної еволюції.
Історія поняття
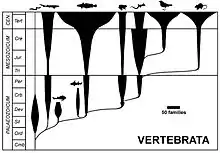
У 1859 р., коли була видана праця Ч. Дарвіна «Походження видів», кількість викопних решток була вкрай малою, науці не були відомі перехідні форми. Дарвін описав відсутність проміжних форм «як найбільш очевидне і найтяжче заперечення, яке може бути проти теорії», але пояснив це крайньою неповнотою геологічного літопису. Він відмічав обмежену кількість доступних колекцій на той час, у той же час описав наявну інформацію про наявні викопні зразки з точки зору еволюції та дії природного добору.[1] Лише двома роками пізніше у 1861 р. був знайдений археоптерикс, який представляв класичну перехідну форму між плазунами і птахами. Його знахіда, стала не тільки підтвердженням теорії Дарвіна, а також знаковим фактом, що підтверджуює реальність існування біологічної еволюції.[2] З того часу була знайдена велика кількість викопних форм, які демонструють, що всі класи хребетних тварин є спорідненими між собою, причому більшість з них — через перехідні форми[3].
Зі збільшенням відомостей про таксономічне різноманіття судинних рослин на початку ХХ ст., почалися дослідження щодо пошуку їхнього можливого предка. У 1917 р. Роберт Кідстон та Вільям Генрі Ленд виявили залишки дуже примітивної рослини біля селища Rhynia в Шотландії.[4] Ця рослина була названа Rhynia. Вона поєднує в собі ознаки зелених водоростей та судинних рослин.
Трактування поняття
Перехідні форми між двома групами організмів не обов'язково є потомками однієї групи і предком іншої. За викопними рештками, як правило, неможливо достеменно встановити чи є певний організм предком іншого. Крім того, ймовірність знайти в палеонтологічному літописі прямого предка певної форми надзвичайно мала. Набагато більша ймовірність виявити відносно близьких родичів цього предка, які подібні з ним за будовою. Тому будь-яка перехідна форма автоматично інтерпретується як бічна гілка еволюції, а не «ділянка філогенетичного стовбура».[5]
Перехідні форми і таксономія
Еволюційна таксономія залишалася домінуючою формою таксономії протягом ХХ ст. Виділення таксонів базується на різноманітних ознаках, внаслідок чого таксони зображують у вигляді гілок розгалуженого еволюційного дерева. Перехідні форми розглядаються як «падаючі» між різними групами в плані анатомії, вони мають суміш характеристик від внутрішньої та зовнішньої клади, що нещодавно розділилася.
Із розвитком кладистики у 1990-х рр. взаємозв'язки зазвичай зображують у вигляді кладограми, яка ілюструє дихотомічне розгалуження еволюційних ліній. Тому в кладистиці перехідні форми розглядаються як більш ранні гілки дерева, де ще розвинулися не всі риси, характерні для раніше відомих нащадків на цій гілці. Такі ранні представники групи зазвичай називають основним таксоном (англ. basal taxa) або сестринським таксоном (англ. sister taxa), залежно від того, чи належить викопний організм до даної клади, чи ні.[6]
Проблеми виявлення та інтерпретації

Відсутність перехідних форм між багатьма групами організмів є предметом критики з боку креаціоністів. Проте далеко не кожна перехідна форма існує у вигляді скам'янілостей через принципову неповноту палеонтологічного літопису. Неповнота викликана особливостями процесу фосилізації, тобто переходу в скам'янілий стан. Для утворення скам'янілості необхідно, щоб організм, який загинув, був похований під великим шаром осадових порід. Через дуже повільну швидкість осадконакопичення на суходолі, сухопутні види рідко переходять у скам'янілий стан та зберігаються. Крім того, рідко вдається виявити види, які мешкають в глибинах океану через рідкісні випадки підняття на поверхню великих масивів дна. Таким чином, більшість відомих викопних (а, відповідно, і перехідних форм) — це або види, що мешкають на мілководді, в морях і річках, або наземні види, які ведуть напівводяний спосіб життя, або мешкають біля берегової лінії. До згаданих вище проблем слід додати надзвичайно малу (в масштабах планети) кількість палеонтологів, які здійснюють розкопки.
Перехідні форми, як правило, не мешкають на великих територіях та не існують упродовж великого часу, інакше вони були б персистентними. Цей факт також знижує ймовірність фосилізації і подальшого виявлення перехідних форм.
Тому ймовірність виявлення проміжних форм надзвичайно мала.
Приклади серед тварин


Найдавнішими представниками земноводних вважають іхтіостегів. Їх вважають перехідною ланкою між кистеперими рибами та земноводними. Незважаючи на те, що у іхтіостегів була п'ятипала кінцівка, адаптована до життя на суші, значну частину життя вони проводили як риби, мали хвостовий плавець, бічну лінію та деякі інші ознаки риб.
Батрахозаврів, що існували в кам'яновугільний і пермський періоди, розглядають як перехідну форму між земноводними і плазунами. Батрахозаври хоч і проводили життя у дорослій стадії на суходолі (подібно до плазунів), були тісно пов'язані з водоймами і зберегли низку ознак, притаманних земноводним, зокрема, відкладання ікри і розвиток личинок у воді, наявність зябер тощо.
Виявлено велику кількість плазунів, які виробили здатність літати, частина з них мала пір'я, тому їх розглядають як перехідні форми між плазунами і птахами. Найбільш відомим є археоптерикс. Він був розміром приблизно з сучасну ворону. Формою тіла, будовою кінцівок та наявністю оперення подібний до сучасних птахів, можливо, літав. Спільним з плазунами була особлива будова таза і ребер, наявність дзьоба з конічними зубами, по три вільні пальці на крилах, подвійноувігнуті хребці, довгий хвіст з 20—21 хребця, кістки були не пневматизовані, грудна кістка без кіля. Інші відомі перехідні форми між плазунами і птахами — протоавіс, конфуціусорніс.
Велика кількість викопних форм звіроподібних плазунів (синапсид, терапсид, пелікозаврів, різних динозаврових тощо.), знайдених у багатьох районах земної кулі, що існували в юрський і крейдовий періоди, поєднують ознаки плазунів і ссавців, розкривають можливі напрями і способи становлення різних груп чотириногих, зокрема ссавців. Наприклад, звіроподібний плазун із групи терапсид — лиценопс (Lycaenops) — за розвитком кісток ротової порожнини, диференціюванням зубів на ікла, різці, за різцеві зуби та низкою інших ознак будови тіла нагадує хижих ссавців, хоча за іншими ознаками та способом життя це були справжні плазуни.
Однією з форм, що збереглася у викопному стані є амбулоцетус Ambulocetus natans («ходячий кит») — перехідна форма між наземними ссавцями і китоподібними, які є вторинноводними формами. Зовні тварина нагадувала дещо середнє між крокодилом і дельфіном. Шкіра мала частково редуковану шерсть. Тварина мала лапи з перетинками; хвіст і кінцівки пристосовані як допоміжні органи пересування у воді.
Приклади серед рослин
Перші наземні рослини з класу риніопсид, родин ринієвих і псилофітових, що жили в силурі — девоні, поєднували ознаки зелених водоростей і примітивних форм вищих рослин. Їх тіло було безлистим, циліндричний осьовий орган — телом у верхній частині дихотомічно розгалуженим на верхівках із спорангіями. Функцію мінерального живлення риніопсид виконували ризоїди.
Викопні форми насінних папоротей, які процвітали наприкінці девону, поєднують в собі ознаки папоротей і голонасінних. Вони утворювали не тільки спори (як папороті), а й насіння (як насінні рослини). Провідна тканина їхніх стебел за будовою нагадує деревину голонасінних (саговників).
Інший попередник насінних рослин був ідентифікований з відкладів середнього девону. Рункарія (Runcaria heinzelinii) існувала близько 20 млн років тому. Це була невелика рослина з радіальною симетрією; мала спорангій, оточений інтегументом та плюскою. Рункарія демонструє шлях еволюції рослин від спорових до насінних.
Перехідні форми в еволюції людини
У наш час знайдена велика кількість викопних рештків, які розкривають еволюційний шлях людини розумної від її людиноподібних предків. До форм, які в більшому чи меншому ступені можна віднести до перехідних, належать: сахелантроп, ардипітеки, австралопітеки (африканський, афарський та інші), людина уміла, людина працююча, людина прямоходяча, людина-попередник, гейдельберзька людина та кроманьйонці.
Серед згаданих форм значної уваги заслуговують австралопітеки. Австралопітек афарський з точки зору еволюції перебуває між сучасними двоногими людьми та їх чотириногими стародавніми предками. Велика кількість рис скелета цього австралопітека чітко відображають двоногість, причому до такого ступеня, що деякі дослідники вважають, що ця властивість виникла задовго до появи австралопітека афарського. Серед загальних рис анатомії, його таз набагато більше подібний до цих кісток у людини, ніж у мавп. Краї клубових кісток коротші та ширші, крижова кістка широка та розташована безпосередньо позаду тазостегнового суглобу. Існує явне свідчення про існування місць кріплення для м'язів-розгиначів коліна, що передбачає вертикальне положення цього організму.[7] У той час, як таз австралопітека не зовсім як у людини (помітно ширший, з орієнтацією краю клубових кісток назовні), ці особливості вказують на принципову перебудову, пов'язану із ходінням на двох ногах. Стегнова кістка утворює кут в напрямку до коліна. Ця риса дозволяє нозі розміщуватися ближче до середньої лінії тіла та є явним свідченням звичного характеру пересування на двох ногах. У наш час людина розумна, орангутани та коати мають такі самі риси. Ноги австралопітека мали великі пальці, що робить практично неможливим захоплення стопою гілок дерев. Крім особливостей локомоції, у австралопітека був також значно більший мозок, ніж у сучасних шимпанзе[8] та його зуби були значно більше подібними до зубів сучасної людини, ніж до мавп.[9]
Філогенетичні ряди
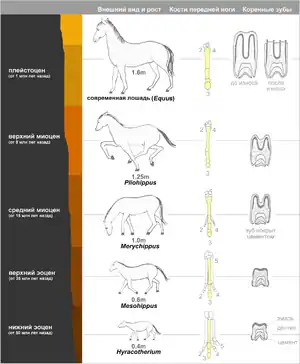
Філогенетичні ряди — ряди викопних форм, пов'язані між собою в процесі еволюції і відбивають поступові зміни їх історичного розвитку.
Були досліджені російським вченим В. О. Ковалевським та англійським Дж. Сімпсоном. Вони показали, що сучасні однопалі копитні походять від стародавніх дрібних всеїдних тварин. Аналіз викопних решток коней допоміг встановити поступовість процесу еволюції в межах цієї групи тварин, зокрема, як змінюючись у часі, викопні форми набували все більшої подібності із сучасними конями. Порівнюючи еоценового еогіпуса із сучасним конем, важко довести їх філогенетичну спорідненість. Проте наявність ряду перехідних форм, які послідовно змінювали одна одну на великих просторах Євразії і Північної Америки, дала змогу відновити філогенетичний ряд коней та встановити напрям їх еволюційних змін. Він складається з ряду наступних форм (у спрощеному вигляді): Phenacodus — Eohippus — Miohippus — Parahippus — Pliohippus — Equus.
Гільгендорф (1866) описав палеонтологічний ряд черевоногих молюсків з міоценових відкладів, що накопичилися протягом двох мільйонів років в озерних відкладах Штейнгеймського басейну (Вюртемберг, Німеччина). Було виявлено в послідовних шарах 29 різноманітних форм, що належали до ряду планорбіс (Planorbis). Стародавні молюски мали черепашку у вигляді спіралі, а пізніші — у вигляді турбоспіралі. Ряд мав два відгалуження. Передбачають, що зміна форми черепашки була викликана підвищенням температури і збільшенням вмісту карбонату кальцію внаслідок гарячих вулканічних джерел.
Таким чином, філогенетичні ряди являють собою історичну послідовність перехідних форм.
У наш час відомі філогенетичні ряди для амонітів (Вааген, 1869), черевоногих молюсків роду живородок (Viviparus) (Neumayr, 1875), носорогів, слонів, верблюдів, непарнокопитних та інших тварин.
Див. також
Посилання
- Дарвин Ч. Происхождение видов путём естественного отбора или сохранение благоприятных рас в борьбе за жизнь. — Перевод с шестого издания (Лондон, 1872). — Санкт-Петербург: Наука, 1991.
- Chiappe, Peter Wellnhofer ; translated by Frank Haase ; foreword by Louis (2009). Archaeopteryx: the icon of evolution (вид. Rev. English). München: F. Pfeil. ISBN 978-3-89937-108-6.
- Prothero, D. (27 лютого 2008). Evolution: What missing link?. New Scientist (2645): 35–40.
- R. Kidston and W.H. Lang (1917) On Old Red Sandstone plants showing structure from the Rhynie chert bed, Aberdeenshire. Part I. Rhynia gwynne-vaughanii, Kidston and Lang. Transactions of the Royal Society of Edinburgh, 5, 761—784.
- Александр Марков. Ископаемые рыбы в очередной раз подтвердили правоту Дарвина (рос.). Элементы.ру. Архів оригіналу за 20 серпня 2011. Процитовано 10 квітня 2014.
- Prothero, Donald R. (2007). Evolution: What the Fossils Say and Why it Matters. Columbia University Press. ISBN 978-0-231-13962-5.
- Lovejoy, C. Owen (1988). Evolution of Human walking. Scientific American 259 (5): 82–89. doi:10.1038/scientificamerican1188-118.
- Australopithecus afarensis. Smithsonian Institution. Процитовано 6 May 2012.
- White, T.D. , Suwa, G., Simpson, S., Asfaw, B. (January 2000). Jaws and teeth of Australopithecus afarensis from Maka, Middle Awash, Ethiopia. American Journal of Physical Anthropology 111 (1): 45–68. PMID 10618588. doi:10.1002/(SICI)1096-8644(200001)111:1<45::AID-AJPA4>3.0.CO;2-I.
- Эволюция конечностей лошади. Государственный Дарвиновский музей. Архів оригіналу за 20 серпня 2011. Процитовано 12 квітня 2014.
Література
- Бровдій В.М. Еволюційне вчення : підручник. — К : ВЦ «Академія», 2013. — 336 с. — ISBN 978-966-580-416-1.
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. — М : Мир, 1992. — 280 с. — ISBN 5-03-001819-0.
