ГКГС класу I
Молекули ГКГС класу I є одним із двох первинних класів молекул головного комплексу гістосумісності (ГКГС, англ. - MHC) (інша — MHC класу II) і знаходяться на клітинній поверхні всіх ядерних клітин в тілах хребетних.[1] [2] Вони також зустрічаються на тромбоцитах, але не на еритроцитах. Їх функція полягає в презентації пептидних фрагментів внутрішньоклітинних білків цитотоксичним Т-клітинам. Така презентація викличе негайну реакцію імунної системи проти певного чужого (невласного) антигена. Оскільки молекули MHC класу I містять пептиди, отримані з цитозольних білків, шлях презентації MHC класу I часто називають цитозольним або ендогенним шляхом . [3]
У людей HLA, що відповідають класу MHCI, є HLA-A, HLA-B і HLA-C.
Функція
Молекули MHC класу I зв'язують пептиди, що утворюються в основному в результаті деградації цитозольних білків в протеасомах. Потім комплекс MHCI-пептид "вставляється" через ендоплазматичний ретикулум у зовнішню плазматичну мембрану клітини. Епітопний пептид зв’язується з позаклітинними частинами молекули MHCI. Таким чином, функція MHCI полягає у презентації внутрішньоклітинних білків цитотоксичним Т-клітинам (ЦТК). Однак MHCI також може презентувати пептиди, отримані з екзогенних білків, у процесі, відомому як "перехресна презентація антигена".
Нормальна клітина на своєму MHCI презентує пептиди нормального обміну білків, і ЦТК не активуються у відповідь на них через центральні та периферичні механізми толерантності. Коли клітина експресує чужорідні білки, наприклад, внаслідок вірусної інфекції, частина MHCI презентуватиме ці пептиди на поверхні клітини. Отже, ЦТК, специфічні для такого комплексу MHC-пептид, будуть його розпізнавати та вбивати клітини, які презентують такі пептиди.
Крім того, сам MHC класу I може служити гальмівним фактором для натуральних кілерів (NK). Зниження експресії поверхневого MHCІ - механізм, який використовується деякими вірусами[4] та деякими пухлинами для ухилення від реакцій ЦТК, активує NK і вони атакують такі клітини.
PirB і зорова пластичність
Парний імуноглобуліноподібний рецептор B (PirB), MHCI-зв’язувальний рецептор, бере участь у регуляції пластичності зору. [5] PirB експресується в центральній нервовій системі і зменшує пластичність очного домінування в критичному періоді розвитку і в дорослому віці.[5] Коли у мутантних мишей функцію PirB було "вимкнено", пластичність очного домінування стала більш вираженою в будь-якому віці.[5] Втрата функції PirB у мутантних мишей також демонструвала підвищену пластичність після монокулярної депривації протягом критичного періоду.[5] Ці результати свідчать про те, що PirB може брати участь у модуляції синаптичної пластичності в зоровій корі .
Структура
Молекули MHC класу I є гетеродимерами, які складаються з двох поліпептидних ланцюгів, α і β2 -мікроглобуліну (B2M). Дві ланцюга пов'язані нековалентно через взаємодію В2M і ділянки α3. Тільки α-ланцюг є поліморфним і кодується геном HLA, тоді як субодиниця B2M не є поліморфною і кодується геном бета-2 мікроглобуліну. Домен α3 заглиблений у плазматичну мембрану і взаємодіє з CD8+ - корецептором Т-клітин. Взаємодія α3-CD8 утримує молекулу MHCI на місці, тоді як Т-клітинний рецептор (TCR) на поверхні ЦТК зв’язує її α1-α2 гетеродимерний ліганд і перевіряє з’єднаний пептид на антигенність. Домени α1 і α2 згинаються, щоб утворити щілину для зв’язування пептидів. Молекули MHC класу I зв’язують пептиди, які мають довжину переважно 8-10 амінокислот (Parham 87), але також повідомлялося про зв’язування довших пептидів.[6]
Синтез
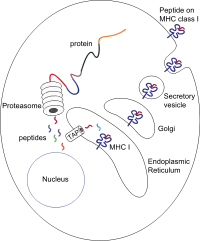
Пептиди в основному генеруються в цитозолі протеасомою. Протеасома — це макромолекула, що складається з 28 субодиниць, половина з яких має протеолітичну активність. Протеасома розкладає внутрішньоклітинні білки до дрібних пептидів, які потім вивільняються в цитозоль. Протеасоми також можуть зшивати окремі фрагменти пептидів (так звані сплайсивані пептиди), утворюючи послідовності, які не є суміжними і, отже, не мають лінійної матриці в геномі. Походження сплайсованих пептидних сегментів може бути від одного білка (цис-сплайсинг) або різних білків (транс-сплайсинг).[7] [8] Пептиди повинні бути переміщені з цитозолю в ЕР, щоб там зустріти молекулу MHC класу I, чий сайт зв’язування пептидів знаходиться в просвіті ЕР.
Транслокація та зв'язування пептиду
Транслокація пептиду з цитозолю в просвіт ЕР здійснюється транспортером, пов’язаним з процесингом антигена (TAP). TAP є членом сімейства транспортерів ABC і являє собою гетеродимерний мультимембранний поліпептид, що складається з TAP1 і TAP2. Дві субодиниці утворюють сайт зв’язування пептиду та два сайти зв’язування АТФ, які звернені до цитозолю. TAP зв’язує пептиди на цитоплазматичній стороні та переміщує їх при споживанні АТФ у просвіт ЕР. Молекула MHC класу I, у свою чергу, зв'язується з пептидами в просвіті ЕР.
У процес зв'язування пептиду залучені кілька інших молекул, які утворюють великий багатовимірний комплекс під назвою на комплекс зв'язування пептиду [9], що складаються з TAP, тапасину, калретікуліну, кальнексіну і ERp57 (PDIA3). Кальнексин стабілізує α-ланцюги MHC класу I до зв’язування β2m. Після повної збірки молекули MHC кальнексин дисоціює. Молекула MHC без зв’язаного пептиду за своєю природою нестабільна і вимагає зв’язування шаперонів кальретикуліну та Erp57. Крім того, тапасин зв’язується з молекулою MHC і служить для зв’язування її з білками TAP і полегшує селекцію пептиду в ітераційному процесі, який називається редагування пептидів,[10] [11] [12] таким чином сприяючи посиленню зв'язування пептидів і колокалізації.
Після того, як пептид зв'язується з молекулою MHC класу I, комплекс дисоціює, і він залишає ЕР через секреторний шлях, щоб досягти поверхні клітини. Транспортування молекул MHC класу I через секреторний шлях включає кілька посттрансляційних модифікацій. Деякі з посттрансляційних модифікацій відбуваються в ЕР і передбачають зміну N-гліканових ділянок білка, за якими слідують великі зміни N-гліканів в апараті Гольджі. N-глікани повністю дозрівають, перш ніж досягти поверхні клітини.
Видалення пептидів
Пептиди, які не зв’язуються з молекулами MHC класу I в просвіті ЕР, видаляються з нього в цитозоль через канал sec61 [13] [14] де вони можуть піддаватися подальшому зменшенню розміру, і можуть бути повернені за допомогою TAP назад в ЕР для зв’язування з молекулою MHC класу I.
Наприклад, спостерігалася взаємодія sec61 з бичачим альбуміном.[15]
Вплив вірусів
Молекули MHC класу I зв'язуються з пептидами, що утворюються в результаті деградації убіквітинованих цитозольних білків у протеасомах. Оскільки віруси індукують клітинну експресію вірусних білків, деякі з цих продуктів позначаються убіквітином для деградації, при цьому отримані пептидні фрагменти потрапляють в ЕР і зв’язуються з молекулами MHCI. Саме таким чином, через презентацію антигена MHCI, заражені вірусом клітини сигналізують Т-клітинам, що в результаті інфекції виробляються аномальні білки.
Внаслідок реакції клітинно-опосередкованого імунітету, інфікована вірусом клітина майже завжди проходить апоптоз, що знижує ризик інфікування сусідніх клітин. Як еволюційна реакція на цей метод імунного нагляду, багато вірусів здатні регулювати або іншим чином перешкоджати презентації молекул MHC класу I на поверхні клітини. На відміну від цитотоксичних Т-лімфоцитів, натуральні кілери (NK) після розпізнавання молекул MHCI на поверхні клітин зазвичай інактивуються. Тому за відсутності молекул MHCI NK-клітини активуються і розпізнають клітину як ненормальну, що свідчить про те, що вона може бути заражена вірусами, які намагаються уникнути імунної відповіді. Деякі види раку людини також викликають зниження експресії MHC I, що дає трансформованим (пухлинним) клітинам ту саму перевагу у виживанні - можливість уникнути нормального імунного нагляду, призначеного для знищення будь-яких інфікованих або трансформованих клітин.[16]
Гени та ізотипи
Еволюційна історія
Гени MHC класу I походять від найближчого спільного предка всіх щелепних хребетних і були знайдені у всіх живих щелепних хребетних, які були вивчені до цього часу.[2] З моменту їх появи у щелепних хребетних це сімейство генів зазнало багатьох різних еволюційних шляхів, оскільки відбувалися видоутворення. Однак існують задокументовані випадки трансвидового поліморфізму в генах MHC класу I, коли конкретний алель в еволюційно спорідненому гені MHC класу I залишається у двох видів, ймовірно, через потужний балансувальний відбір через дію патогенів, які можуть інфікувати обидва види.[17] Гіпотеза "еволюція народження і смерті" є одним із механістичних пояснень розміру сімейства генів класу MHC I.
"Еволюція народження і смерті" генів MHC класу I
Гіпотеза "еволюції народження і смерті" стверджує, що явища дуплікації генів призводять до того, що геном містить кілька копій гена, які потім можуть зазнавати окремих еволюційних процесів. Іноді ці процеси призводять до псевдогенізації (смерті) однієї копії гена, хоча іноді цей процес призводить до появи двох нових генів з різною функцією.[18] Ймовірно, що локуси MHC класу Ib людини (HLA-E, -F і -G), а також псевдогени MHC класу I виникли з локусів MHC класу Ia (HLA-A, -B і -C) у цьому процесі "народження і смерті".[19]
Посилання
- The MHC class I antigen presentation pathway: strategies for viral immune evasion. Immunology 110 (2): 163–9. October 2003. PMC 1783040. PMID 14511229. doi:10.1046/j.1365-2567.2003.01738.x. Проігноровано невідомий параметр
|vauthors=(довідка) - Comparative genomic analysis of the MHC: the evolution of class I duplication blocks, diversity and complexity from shark to man. Immunological Reviews 190: 95–122. December 2002. PMID 12493009. doi:10.1034/j.1600-065x.2002.19008.x. Проігноровано невідомий параметр
|vauthors=(довідка) - http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/H/HLA.html#Class_I_Histocompatibility_Molecules Kimball's Biology Pages, Histocompatibility Molecules
- MHC class I antigen presentation: learning from viral evasion strategies. Nature Reviews. Immunology 9 (7): 503–13. July 2009. PMID 19498380. doi:10.1038/nri2575. Проігноровано невідомий параметр
|vauthors=(довідка) - PirB restricts ocular-dominance plasticity in visual cortex. Science 313 (5794): 1795–800. September 2006. Bibcode:2006Sci...313.1795S. PMID 16917027. doi:10.1126/science.1128232. Проігноровано невідомий параметр
|vauthors=(довідка) - Have we cut ourselves too short in mapping CTL epitopes?. Trends in Immunology 27 (1): 11–6. January 2006. PMID 16297661. doi:10.1016/j.it.2005.11.001. Проігноровано невідомий параметр
|vauthors=(довідка) - Faridi, Pouya; Li, Chen; Ramarathinam, Sri H.; Vivian, Julian P.; Illing, Patricia T.; Mifsud, Nicole A.; Ayala, Rochelle; Song, Jiangning та ін. (12 жовтня 2018). A subset of HLA-I peptides are not genomically templated: Evidence for cis- and trans-spliced peptide ligands. Science Immunology 3 (28): eaar3947. PMID 30315122. doi:10.1126/sciimmunol.aar3947. Проігноровано невідомий параметр
|doi-access=(довідка); - Liepe, Juliane; Marino, Fabio; Sidney, John; Jeko, Anita; Bunting, Daniel E.; Sette, Alessandro; Kloetzel, Peter M.; Stumpf, Michael P. H. та ін. (21 жовтня 2016). A large fraction of HLA class I ligands are proteasome-generated spliced peptides. Science 354 (6310): 354–358. Bibcode:2016Sci...354..354L. PMID 27846572. doi:10.1126/science.aaf4384. Проігноровано невідомий параметр
|hdl-access=(довідка); - Structure of the human MHC-I peptide-loading complex. Nature 551 (7681): 525–528. November 2017. Bibcode:2017Natur.551..525B. PMID 29107940. doi:10.1038/nature24627. Проігноровано невідомий параметр
|vauthors=(довідка) - Tapasin enhances MHC class I peptide presentation according to peptide half-life. Proceedings of the National Academy of Sciences of the United States of America 101 (32): 11737–42. August 2004. Bibcode:2004PNAS..10111737H. PMC 511045. PMID 15286279. doi:10.1073/pnas.0306294101. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Selective loading of high-affinity peptides onto major histocompatibility complex class I molecules by the tapasin-ERp57 heterodimer. Nature Immunology 8 (8): 873–81. August 2007. PMID 17603487. doi:10.1038/ni1485. Проігноровано невідомий параметр
|vauthors=(довідка) - Tapasin shapes immunodominance hierarchies according to the kinetic stability of peptide-MHC class I complexes. European Journal of Immunology 38 (2): 364–9. February 2008. PMID 18196518. doi:10.1002/eji.200737832. Проігноровано невідомий параметр
|vauthors=(довідка) - Export of antigenic peptides from the endoplasmic reticulum intersects with retrograde protein translocation through the Sec61p channel. Immunity 13 (1): 117–27. July 2000. PMID 10933400. doi:10.1016/S1074-7613(00)00013-3. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Retrotranslocation of MHC class I heavy chain from the endoplasmic reticulum to the cytosol is dependent on ATP supply to the ER lumen. Molecular Immunology 40 (10): 733–41. January 2004. PMID 14644099. doi:10.1016/j.molimm.2003.08.008. Проігноровано невідомий параметр
|vauthors=(довідка) - Exogenous antigens are processed through the endoplasmic reticulum-associated degradation (ERAD) in cross-presentation by dendritic cells. International Immunology 17 (1): 45–53. January 2005. PMID 15546887. doi:10.1093/intimm/dxh184. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Activation of CXCR4 triggers ubiquitination and down-regulation of major histocompatibility complex class I (MHC-I) on epithelioid carcinoma HeLa cells. The Journal of Biological Chemistry 283 (7): 3951–9. February 2008. PMID 18083706. doi:10.1074/jbc.m706848200. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Trans-species polymorphism in humans and the great apes is generally maintained by balancing selection that modulates the host immune response. Human Genomics 9: 21. September 2015. PMC 4559023. PMID 26337052. doi:10.1186/s40246-015-0043-1. Проігноровано невідомий параметр
|vauthors=(довідка) - Concerted and birth-and-death evolution of multigene families. Annual Review of Genetics 39 (1): 121–52. 14 листопада 2005. PMC 1464479. PMID 16285855. doi:10.1146/annurev.genet.39.073003.112240. Проігноровано невідомий параметр
|vauthors=(довідка) - Origin and evolution of HLA class I pseudogenes. Molecular Biology and Evolution 12 (2): 247–58. March 1995. PMID 7700152. doi:10.1093/oxfordjournals.molbev.a040201. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка)