Гідроїдні
Гідро́їдні — клас кнідарій, чий життєвий цикл включає медузу з характеристичною ознакою — велумом, та поліпа, що, на відміну від інших кнідарій, не має внутрішніх перетинок (септ) та вираженої глотки.
? Гідроїдні | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Solanderia ericopsis | ||||||||||||
| Біологічна класифікація | ||||||||||||
| ||||||||||||
|
Trachylinae Leptolinae | ||||||||||||
Посилання | ||||||||||||
| ||||||||||||
Загальна характеристика
Життєвий цикл може не мати стадії поліпу або медузи, але обов'язково включає личинку-планулу. Спосіб життя може бути поодиноким (гідра) або колоніальним (обелія), у більшості видів у поліпових стадій формуються колонії; зустрічаються колонії, в яких інтегровані одночасно і поліпи, і медузи (ряд Сифонофори).
Еволюція
Викопні рештки гідроїдних відомі з докембрію; втім, з огляду на малу кількість твердих скелетних структур ці рештки є досить нечисленними та фрагментарними. Останні дослідження будови медузарної нодули (спеціальна структура на поліпі, що утворює молодих медуз шляхом специфічного брунькування) дали результати, що свідчать про наявність у гідроїдних трьох зародкових листків (тобто гідроїдні є тришаровими). Підпарасолькова порожнина гідромедуз та шар смугастого м'яза, що її вистилає, утворюються з морфологічної структури, що дуже нагадує шизоцель: в цьому випадку між ектодермою та ентодермою формується третій шар (аналог мезодерми), який перетворюється на порожнину. Таким чином, підпарасолькова порожнина є, фактично, целомом, який згодом, після формування веларного отвору, стає відкритим назовні.
Також зараз наявні молекулярно-біологічні дані, які показують, що гени, котрі кодують формування структур мезодерми у двосторонньо-симетричних тварин (Bilateria) присутні також і у гідроїдів. Таким чином, стадія поліпа у гідроїдів має два зародкових листки (тобто є двошаровою), а стадія медузи — три зародкових листки (тобто є тришаровою). Якщо ці дані будуть підтверджені доказами ще й з інших джерел, це буде означати, що перехід від двошарових до тришарових організмів відбувається щоразу при відбрунькуванні медузи від поліпу, і, таким чином, буде вирішене одне з найбільших питань еволюції тварин — яким чином відбувся перехід від Diploblasta (тварин з двома зародковими листками) до Tryploblasta (тварин з трьома зародковими листками).
Систематика
Гідроїдні відомі систематикам з самого початку існування зоології як такої; велика кількість видів описані Карлом Ліннеєм ще у вісімнадцятому сторіччі.
Систематика гідроїдних досить складна, що викликано браком палеонтологічних відомостей, на основі яких можна виявити родинні зв'язки всередині таксона. Зараз існує кілька загальних варіантів класифікації; в даній статті класифікація побудована за принципами, загалом викладеними на сайті The Hydrozoa Directory, і у великій мірі базується на результатах молекулярно-біологічних досліджень останніх років. Згідно з результатами згаданих досліджень, клас Гідроїдні наочно розподіляється на дві групи рядів, що отримали статус підкласів: Trachylinae та Leptolinae (останній в інших джерелах називається також Hydroidolina та Hydoidomedusae).
В дану класифікаційну систему не включений внутрішньоклітинний паразит поліподіум (Polypodium hydriforme), з огляду на його високу спеціалізацію та неясні родинні зв'язки і походження; згідно з деякими відомостями (Siddall et al., 1995) навіть його позиціонування серед кнідарій не є беззаперечним.
Гідроїдні є таксоном-космополітом, тобто таким, що розповсюджений в усьому світі. Вони зустрічаються як у прісній, так і у солоній воді.
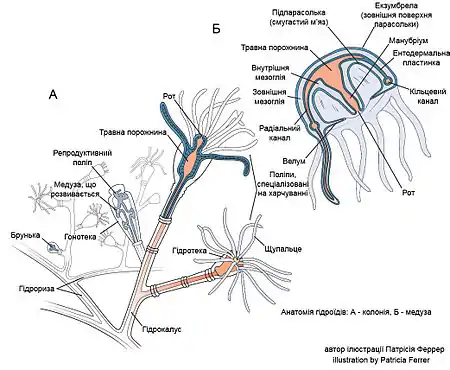
Спосіб життя
Медузові стадії життєвого циклу Гідроїдних, а також поліпові стадії у сифонофор, є в більшості планктонними організмами. Вони зустрічаються сезонно, часто у великих скупченнях, що переносяться течіями. Деякі медузи та сифонофори, втім, є бентосними. Поліпові стадії звичайно належать до бентосу, і ведуть осілий спосіб життя, але і тут є винятки: відомо кілька планктонних гідроїдних поліпів. Зокрема, саме планктонним вільноплаваючим поліпом є так звана медуза-вітрильник (Velella velella). Широко відомий португальський кораблик також є вільноплаваючою колонією, сформованою із спеціалізованих гідроїдних поліпів.
Гідроїдні зустрічаються в усіх типах водних екосистем, від наземних печер до глибоководних океанських жолобів, від озер та ставків до скельних узбереж та країв антарктичних шельфових льодовиків, а також у пустотах між піщинками в лінії прибою. Поліпові стадії багатьох видів часто прикріплюються до специфічного виду субстрату, звичайно інших організмів: риб, покривників, поліхет, мшанок, молюсків, ракоподібних, губок, кишковопорожнинних, водоростей, голкошкірих, і т. ін., де вони вступають з хазяїном у відношення, що можуть характеризуватись як коменсалізм, мутуалізм, або паразитизм.
Більшість гідроїдних є хижаками, що використовують особливості свого способу життя для ловлі здобичі. Планктонні стадії, що транспортуються течією, часто здатні також і до активного руху в пошуках їжі. Розташування прикріплених форм визначається тим, де осяде планула. Колонії поліпів звичайно виникають у місцях, де забезпечений постійний потік води, що збільшує надходження потенційної їжі.
Поведінка
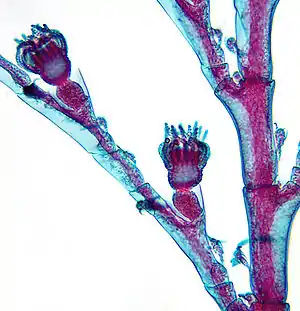
Медузи ведуть строго індивідуальний спосіб життя; вони можуть бути зігнаними вітром та течією у великі скупчення, але досі у них не реєструвалося ніяких форм соціальної поведінки. Колонії ж гідроїдних поліпів, особливо поліморфні, за рівнем спеціалізації окремих поліпів та узгодженості їхніх дій можуть бути порівняні з єдиним організмом. Поліпи в колонії звичайно є нащадками єдиної планули, і, таким чином, є поєднаними клонами з ідентичним генотипом. Тим не менше, у деяких видів колонії можуть змішувати свої тканини або потомки кількох планул — формувати єдину колонію. В цих випадках різні особини поліпів знаходяться в такому тісному взаємозв'язку, що формують (на функціональному, але не на генетичному рівні) єдиний організм, що є, вірогідно, однією з найтісніших форм соціальної організації.
Більшість гідроїдних є роздільностатевими. Запліднення зазвичай внутрішнє, без копуляції. Чоловічі особини викидають у воду сперму, активно плаваючи повз яйця, що прикріплені до материнського організму (медузи або поліпу) або викинуті у воду жіночою особиною. Гідроїдні — перші організми, у яких було продемонстровано наявність сперматозоїдних атракторів (речовин, що принаджують сперматозоїди при їхньому вільному русі), які забезпечують видоспецифічну атракцію сперматозоїдів до яєць.
Члени однієї колонії поліпів (зооїди) вдаються до координованої поведінки, що вимагає деякої комунікації між ними. У таких видів як, наприклад, Thecocodium brieni, дактилозооїди ловлять здобич щупальцями, тоді як гастрозооїди, після захоплення здобичі витягуються до дактилозооїдів, виймають здобич з їхніх щупалець та ковтають її. Такий поділ праці, що передбачає розвинену координацію, для поліморфних колоній є досить звичайним.
Очевидно, що планктонні організми не можуть виявляти яскравої територіальної поведінки; але, як показав ряд досліджень, вільноплаваючі стадії в життєвому циклі гідроїдних активно уникають надто щільного скупчення особин свого виду при харчуванні. Територіальна поведінка є яскраво вираженою серед бентосних організмів, де конкуренція за придатні місця розташування є зазвичай високою. Таким чином, висока концентрація жалячих дактилозооїдів на периферії колонії (у колоніальних видів) є захисним пристосуванням, спрямованим на обмеження росту оточуючих тварин. В цих же колоніях гастрозооїди здатні до поїдання осідаючих планул інших видів, що при розвитку можуть скласти конкуренцію.
І медузи, і поліпи в голодному стані безперервно рухаються в пошуку їжі; коли ж травна порожнина (коелентерон) заповнюється, щупальця звичайно скорочуються та підтягуються до тіла, що забезпечує деякий ступінь контролю за раціональною витратою жалячих клітин (кнідоцитів). Харчова поведінка медуз багатьох видів зумовлює їхні періодичні вертикальні міграції.
Харчування


Основним харчовим ресурсом гідроїдних є планктон — зокрема, дрібні ракоподібні. В лабораторних умовах основою харчування гідроїдів є, звичайно, артемія. Медузи гідроїдів є, в більшості, строгими хижаками, і, у випадку харчування ікрою та личинками риб можуть розглядатись як вершина харчової піраміди.
Раціон поліпів є різноманітнішим; деякі види мають симбіотичних одноклітинних водоростей, і часом харчуються винятково за рахунок поживних речовин, що їх ті постачають при фотосинтезі. Таким чином, ці види можуть вважатись функціонально фотосинтетичними тваринами.
Форми вилову здобичі медузами варіюють від пасивного ширяння в товщі води з нерухомими щупальцями, на які може наштовхуватись їстівний планктон, до активного плавання в пошуках об'єктів харчування. Поліпи здатні до видовження щупалець та рухів ними для вловлювання пропливаючої здобичі, але також можуть вдаватись до цілеспрямованого полювання, котре забезпечують наявні (не у всіх видів) чутливі органи, що сигналізують про наближення здобичі.
Симбіотичні види можуть бути дуже спеціалізованими в харчовій поведінці: так, Halocoryne epizoica харчується щупальцями мшанок, а Polypodium hydrifirme — ікринками осетрових риб, виїдаючи їх зсередини, і, таким чином, є єдиною багатоклітинною твариною — внутрішньоклітинним паразитом.
Основним знаряддям полювання у гідроїдних є кнідоцити. Гідроїдам притаманний найширший спектр типів цих жалячих клітин серед усіх кнідарій.
З екологічної точки зору, гідроїдні медузи, що харчуються ікрою риб, є найнебезпечнішими для них хижаками; а здатність поліпів до харчування практично будь-якими личинками риб та ракоподібних включає їх як важливу ланку в життєвий цикл величезної кількості видів. Таким чином, екологічне значення харчової спеціалізації гідроїдних є дуже великим.
Розмноження


У гідроїдних не виявлено жодних ознак спеціальної шлюбної поведінки.
Яйця зберігаються у статевих залозах (гонофорах) жіночих особин. Відповідно до виду, яйця можуть бути дрібними й чисельними, або великими та нечисельними, аж до одного великого яйця на гонофор.
У видів, що не мають в життєвому циклі поліпової стадії, безстатеве розмноження відсутнє, або наявне у невеликій мірі на стадії личинкової медузи (що відома як актинула в ряді Narcomedusae), тобто кожне запліднене яйце дає початок одній або невеликій кількості дорослих особин. В підкласі Leptolinae (інша назва Hydromedusae) широко розповсюджене безстатеве розмноження на стадії поліпа, яке вважають аналогом поліембріонії, або личинкового розмноження, характерного для паразитичних тварин (таких, наприклад, як трематоди). Кожне запліднення яйця, в такому випадку, призводить до появи однієї планули, з якої розвивається колонія поліпів, котра, в свою чергу, продукує багато дорослих медуз. Позаяк життєвий цикл поліпової колонії може продовжуватись багато років, кожна планула, таким чином, стає попередником сотень тисяч медуз.
Планула гідроїдних є, фактично, ембріоном, а не личинкою, з огляду на свою вкрай просту будову (фактично це гаструла). Планула гідроїдних може бути пустотілою (тобто целобластулою) або без внутрішньої порожнини, (тобто стереогаструлою); звичайно видам, які мають медузу в життєвому циклі, притаманна пустотіла планула, яка проводить частину свого життя в товщі води, плаваючи за допомогою війок епітелію. Види ж, у яких медузоїдна стадія у життєвому циклі відсутня, звичайно продукують планули без внутрішньої порожнини, що відразу осідають на дно поряд з материнським організмом (або колонією). У випадку наявності медузи в життєвому циклі саме це покоління є «статевим», тобто здатним до статевого розмноження. Поліпове покоління, таким чином, є спеціалізованою та багаторічною личинкою, що продукує велику кількість статевих особин протягом свого існування. У багатьох видів, втім, медузоїдна стадія може бути частково або навіть повністю редукована, і в такому випадку личинка (поліпова стадія) завдяки педоморфозу стає статевозрілою особиною. Майже для половини видів підкласу Leptolinae характерна редукована або відсутня медузоїдна стадія; таким чином, ця група є таксоном з найпоширенішим педоморфозом серед усіх тварин.
Деякі медузи (наприклад, рід Eleutheria) мають спеціальні виводкові кишені, де утримують дрібних молодих медуз. Також для деяких гідроїдів характерні гонотеки з виводковими камерами, де деякий час утримуються планули.
Багато видів гідроїдних є строго сезонними, являючись активними тільки протягом деякого проміжку часу. Медузи можуть спостерігатись протягом тижнів або місяців, після чого повністю зникають з водної товщі, і вигляд протягом всього іншого проміжку року представлений відповідними поліпами в бентосі. Поліпові колонії, в свою чергу, можуть регресувати до перечікуючої протягом довгого періоду гідрорізи, реактивуючись при поверненні сприятливих умов існування. Планула може інцистуватись і перечікувати несприятливі умови аналогічно гідроризі, будучі вкритою захисною хітиновою оболонкою.
Охоронний статус
У Червоному Списку МСОП немає жодного виду класу Гідроїдні. Для більшості видів точні межі ареалу та чисельність є невідомими. Велика кількість видів вважається ендемічними просто з тієї причини, що їхнім цілеспрямованим пошуком не займались за межами ареалу первинного відкриття.
У регіональних та національних Червоних книгах присутні такі представники гідроїдних, як кальцієфіковані, коралоподібні родини Milliporidae та Stylasteridae, котрі також занесені до списку видів CITES. Вказані родини є об'єктами торгівлі, поряд з деякими іншими гідроїдами (відомими у акваторії Північного моря як «білі водорості»). Зменшення їхньої чисельності відбувається, в основному, завдяки руйнуванню життєвого середовища.
До Червоної Книги України занесені два види цього класу: оліндіас несподіваний (Olindias inexpectata) та меризія азовська (Moerisia maeotica).
Значення для людини
Відомий трактат Тремблі, де описані трансформації гідроїдів роду Hydra, надихнув Мері Шеллі на написання роману «Франкенштейн»; сучасний композитор Франк Заппа написав пісню про гідромедузу, названу зоологами на його честь — Phialella zappai. Але, звичайно, гідроїди не привертають до себе особливої уваги людей.
«Білі водорості» (колонії поліпів родів Hydrallmania та Sertullaria) раніше використовувались як декоративні прикраси, доки популяції цих гідроїдів не стали катастрофічно скорочуватись. Деякі гідроїдні слугують лабораторними тваринами: класичним прикладом є поліпи роду Hydra, що, поряд з науковими дослідженнями, використовуються у шкільному навчанні в багатьох країнах світу; але гідри не є єдиним прикладом такого використання: також в науковій роботі широко використовується Aequorea victoria (для отримання білка-маркера екворина), та види з родів Hydractinia, Laomedea і Tubularia.
Гідроїди є важливими членами так званих «забруднюючих співтовариств», що населяють водопостачальні мережі електростанцій, утруднюючи їхню роботу, а також погіршують ходові характеристики кораблів, заселяючи їхню підводну обшивку. Деякі види є шкідниками аквакультур, завдяки своєму харчуванню личинками цінних видів, або конкуруванню з ними за їжу. Гідроїд Polypodium hydriforme є шкідником осетрової промисловості, позаяк він є внутрішнім паразитом ікри осетрів.
Медузи деяких видів можуть завдавати серйозні опіки людині; також ця небезпека існує і при контакті з поліповими колоніями таких видів, таких, наприклад, як вогняні корали (Millepora). При пересуванні великими стадами навіть дрібні медузи, такі як представники роду Cytia, можуть завдавати відчутні опіки плавцям.
Але найбільшої шкоди людині завдає саме харчування деяких медуз (наприклад Aeroquorea victoria) та вільноплаваючих колоній поліпів (таких як Cytia gracilis) личинками та ікрою промислових риб.
Література
- Bouillon, J. 1985. Essai de classification des hydropolypes-hydroméduses (Hydrozoa-Cnidaria). Indo Malayan Zoology 2: 29-243.
- Bouillon, J. 1994. Les Hydrozoaires. In Traité de Zoologie, vol. 3(2) (ed. P. Grassé & J. Doumeng), pp. 29-416. Paris: Masson.
- Bouillon, J., & Boero, F. 2000b. Synopsis of the families and genera of the Hydromedusae of the world, with a list of the worldwide species. Thalassia Salentina 24: 47-296.
- Brusca, R. C., and G. J. Brusca. Invertebrates. 1990. Sinauer Associates, Sunderland, MA.
- Cornelius, P. F. S. 1995. North-west European thecate hydroids and their medusae. Synopses of the British Fauna New Series 50, 2 volumes.
- Collins, A. G. 2002. Phylogeny of Medusozoa and the evolution of cnidarian life cycles. Journal of Evolutionary Biology 15: 418–432.
- Gili, J. M. & Hughes, R. G. 1995. The ecology of marine benthic hydroids. Oceanography And Marine Biology An Annual Review 33: 351–426.
- Grzimek's Animal Life Encyclopedia, Edition 2, Volume 01; Thomson Gale, 2003.
- Hyman, L. H. 1940. The Invertebrates: Protozoa through Ctenophora. New York and London: McGraw-Hill.
- Marques, A. C. 2001. Simplifying hydrozoan classification: inapproriateness of the group Hydroidomedusae in a phylogenetic context. Contributions to Zoology 70: 175–179.
- Millard, N. A. H. 1975. Monograph on the Hydroida of southern Africa. Annals of the South African Museum 68: 1-513.
- Schuchert, P. 1993. Phylogenetic analysis of the Cnidaria. Zeitschrift für Zoologische Systematik und Evolutionsforschung 31: 161–173.
- Siddall, M. E., Martin, D.S., Bridge, D., Desser, S. S., & Cone, D. K. 1995. The demise of a phylum of protists: phylogeny of Myxozoa and other parasitic Cnidaria.Journal of Parasitology 81: 961–967.
- Werner, B. 1984. 4. Stamm Cnidaria, Nesseltiere. In Wirbellose Tiere (ed. H.-E. Gruner), pp. 10-305. Stuttgart: Gustav Fischer.
Посилання
- Актинула // ВУЕ
- J. Bouillon, M.D. Medel, F. Pagès, J.M. Gili, F. Boero and C. Gravili. 2004. Fauna of the Mediterranean Hydrozoa. Scientia Marina, 68 (Suppl. 2).
- Hydroids from Reunion Island and Indian Ocean
- https://web.archive.org/web/20051025163703/http://zygote.swarthmore.edu/intro6.html
- Puget Sound Online
- Aquascope
- The Hydrozoa Directory
|
|
Ця стаття належить до вибраних статей Української Вікіпедії. |