Молюски
Молю́ски, або м'якуни́ (Mollusca) — тип первинноротих двобічно-симетричних целомічних тварин зі спіральним дробленням зиготи.
? МолюскиЧас існування: Ранній кембрій — наш час | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Виноградний равлик — один з найпоширеніших та найбільших молюсків України | ||||||||||||
| Біологічна класифікація | ||||||||||||
| ||||||||||||
Класи | ||||||||||||
| ||||||||||||
Посилання | ||||||||||||
| ||||||||||||
М'якуни — другий за чисельністю видів тип тварин після Членистоногих. Наразі відомо приблизно 130 тисяч сучасних видів молюсків[1][2], проте науковці припускають існування до 200 тисяч видів[1][3]. Крім того, відомо від 35[2] до 70[1] тисяч викопних видів м'якунів. Приблизно 80 % видів належать до класу Черевоногих. В Україні налічують орієнтовно 500—650 видів молюсків.
Здебільшого молюски є водяними тваринами, що мешкають у морських та прісних водоймах, меншість м'якунів наземні[⇨]. Здебільшого молюски є вільноживучими організмами, проте деякі з них пристосувалися до паразитичного способу життя.
Молюски дуже різноманітні за розмірами, анатомічною будовою та поведінкою. Дорослі особини найменших видів (належать до черевоногих та двостулкових молюсків) сягають завдовжки лише 0,5 мм[4][5][6], тоді як найбільші — велетенські кальмари з класу Головоногих, зокрема Mesonychoteuthis hamiltoni, які можуть досягати 10 м у довжину та 495 кг ваги[7], є найбільшими безхребетними тваринами. Однак переважна більшість м'якунів має розміри в кілька сантиметрів[4].
Одна з характерних особливостей молюсків — мінералізована черепашка (мушля), форма та будова якої відрізняється у представників різних класів. У більшості сучасних головоногих мушля редукована. Характерним для молюсків є також особливий ротовий орган — радула. У двостулкових радула, як і вся голова, повністю редукована. Пересувається більшість молюсків за допомогою ноги, яка в головоногих перетворилася на щупальця. Головоногі молюски, а саме кальмари, каракатиці та восьминоги, мають одну з найрозвиненіших нервових систем серед усіх безхребетних[⇨].
Життєві цикли молюсків також досить різноманітні. Наземні м'якуни та представники класу Головоногих розвиваються без проміжних стадій, інші ж мають різні личинкові форми[⇨].
Появу перших молюсків наразі датують початком кембрійського періоду палеозойської ери (приблизно 540 млн років тому)[⇨]. Класифікація, філогенетика та навіть монофілія молюсків як типу наразі остаточно не встановлені, проте найчастіше тип поділяють на 8 сучасних та 2 викопних класи[⇨].
Багато видів молюсків, переважно наземних та прісноводних, опинилися перед загрозою вимирання через діяльність людини та потребують охорони. Наразі 1032 сучасні види цієї групи вже відмічені як імовірно вимерлі[8], що становить більше половини всіх відомих сучасних вимирань видів[⇨].
Певні види молюсків є важливим джерелом харчування для людини, а також джерелом цінних матеріалів, зокрема перламутру, перлів, пурпуру та висону, з яких виготовляють коштовні прикраси та інші предмети розкошів[⇨]. Водночас деякі м'якуни є сільськогосподарськими шкідниками, а деякі можуть безпосередньо загрожувати здоров'ю та життю людини[⇨].
У популярній культурі равлики є символом неквапливості, а велетенські головоногі надихали давніх мореплавців та сучасних авторів на розповіді про морських чудовиськ[⇨].
Вивчає молюсків наука малакологія.
Етимологія
Назва «молюск» походить від латинського molluscus — «м'який»[9][10][11]. Вивчає молюсків наука малакологія, а їхні мушлі — конхологія[12] або конхіологія[13].
Будова та фізіологія

Особливості зовнішньої та внутрішньої будови тіла різних груп молюсків є досить значними, що дуже ускладнює виявлення синапоморфій (ознак, спільних для всіх представників даної групи організмів, але відсутніх у їхніх предків), які об'єднали б усі сучасні класи м'якунів[15][16]. Найзагальнішими характерними рисами молюсків можна вважати відсутність справжньої сегментації та двосторонню симетрію[17]. Однак у різних груп м'якунів тіло набуває асиметричності через зміщення або нерівномірний ріст різних органів. Найяскравіше асиметрія проявляється в черевоногих молюсків унаслідок торсійного процесу та виникнення турбоспіральної мушлі. Спільними ознаками є також наявність мантії та мантійної порожнини, яка виконує дихальну та видільну функції, а також будова нервової системи.
Тіло
Тіло м'якунів не має ознак справжньої сегментації[18], попри те, що деякі органи (наприклад, зябра хітонів та моноплакофор) можуть мати сегментовану будову. Зазвичай тіло молюсків складається з чотирьох відділів: голови, ноги, нутрощевого мішка (вісцеральної маси) та мантії[19]. У представників класу Ямкохвостих відсутня нога, а двостулкові молюски вторинно втратили голову.
Нога являє собою м'язистий непарний виріст черевної стінки тіла і слугує переважно для пересування[20], проте у представників різних класів може виконувати інші функції[21]. У нозі розташовані два органи рівноваги (статоцисти). Нога черевоногих виділяє слиз, який змащує поверхню й полегшує пересування. У молюсків, що мають черепашку розташовану лише на верхній частині тіла (як у морського блюдечка), нога прикріпляє тварину до твердої поверхні завдяки дії вертикальних м'язів, які притискають тіло до неї. В інших м'якунів вертикальні м'язи втягують ногу та інші м'які частини тіла всередину мушлі[22]. Двостулкові мають ногу пристосовану для закопування у ґрунт[21], хоча деякі, наприклад, мідії, її втратили. У головоногих молюсків нога перетворилася на щупальця[23] і бере участь у реактивному русі[21].
Тулуб містить усі основні внутрішні органи. У представників групи Conchifera він сильно розростається на дорсальний бік у процесі ембріонального розвитку, внаслідок чого формується вісцеральна маса.
Від основи тулуба відходить мантія — складка стінки тіла, вкрита епідермісом, що утворює мантійну порожнину, яка сполучається із зовнішнім середовищем. У мантійній порожнині міститься так званий мантійний комплекс органів: вивідні шляхи статевої, травної та видільної систем, ктенидії, осфрадії та гіпобранхіальна залоза. Крім того, до мантійного комплексу органів належать нирки та перикард, розташовані поруч із мантійною порожниною. У ранніх молюсків мантійна порожнина була розташована ближче до задньої частини тіла, проте в сучасних групах її розташування широко варіює. У двостулкових молюсків усі м'які частини тіла лежать у межах мантійної порожнини[24].
Покриви

Вважається, що в гіпотетичного предка молюсків покрив був представлений протоперінотумом — кутикулою з арагонітовими спікулами. Подібна будова покрову притаманна представникам класів Caudofoveata і Solenogastres. Проте у всіх класів молюсків, окрім Caudofoveata, з'являється війкова плазівна поверхня — нога (за цією ознакою їх об'єднують у групу Adenopoda). У Solenogastres нога представлена педальною борозною. Панцирні також мають кутикулярний покрив, але тільки на латеральних поверхнях — перинатальних складках. Дорсальна ж поверхня прикрита вісьмома пластинками черепашки.
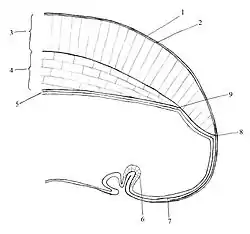
Мінерально-органічна черепашка є характерною ознакою молюсків, яка в типових випадках вкриває все тіло молюска і виконує захисну функцію. Черепашка може бути суцільною чи складатися з кількох частин. У деяких груп черепашка повністю чи частково редукована. Як правило, черепашка молюсків складається з трьох шарів: зовнішнього конхіолінового (периостракума), складеного з органічної речовини — конхіоліну; середнього призматичного (остракума), до складу якого входить карбонат кальцію у вигляді призматичних кристалів, розташованих перпендикулярно до поверхні черепашки; внутрішнього перламутрового (гіпостракума), що складається із тонких пластинчатих кристалів карбонату кальцію, розташованих паралельно до поверхні черепашки. Черепашка утворюється завдяки секреторній діяльності епітелію краю мантії.
У представників групи Conchifera, що включає класи Черевоногі, Головоногі, Двостулкові, Лопатоногі та Моноплакофори, кутикулярний покрив відсутній, а черепашка складається з однієї пластинки. Під черепашкою лежить мантія — складка шкіри, яка вільно звисає по боках тулуба й огортає його основу. Між тулубом і мантією залишається мантійна порожнина, у якій містяться органи дихання — зябра або легені, слинні залози, органи хімічного чуття.
Вторинна порожнина тіла
Вторинна порожнина тіла в молюсків являє собою справжній, але значно редукований целом (заповнену рідиною порожнину тіла, вкриту серозною мембраною). Основний об'єм целому в них представлений міксоцелем, що є частиною незамкненої кровоносної системи. Целомічні мішки в молюсків представлені перикардом (порожнина серцевої сумки) та порожниною гонад. Разом вони утворюють гоноперикардіальну систему. Основною порожниною організму молюсків є гемоцель, через який циркулює кров та целомічна рідина, проміжки між органами частково заповнені паренхімою. Нирки фактично є целомодуктами[25], пов'язаними з перикардом. Передсердя виконують частину функцій видільної системи, відфільтровуючи відходи метаболізму з крові та скидаючи їх у целом вже у вигляді сечі[22]. Целомодукти, що відкриваються в порожнину гонад — статеві протоки (гонодукти)[26].
Травна система

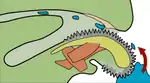
Травна система м'якунів починається з ротового отвору, що веде в ротову порожнину, у яку зазвичай відкриваються слинні залози. Травна система складається з глотки, стравоходу, шлунку, середньої та задньої кишок (ректум). Є також травна залоза — печінка, яка бере участь у перетравлюванні, всмоктуванні та накопиченні поживних речовин (клітини печінки молюсків здатні до фагоцитозу). У головоногих є також підшлункова залоза (у решти молюсків її функції виконує травна залоза).
У більшості видів у глотці є радула («тертушка»)[27] — спеціальний апарат для подрібнення їжі. Радула вкрита хітиновими зубцями, що змінюються в міру їх зношення. Головною функцією радули є зішкрібання бактерій та водоростей з каміння та інших поверхонь. Радула пов'язана з одонтофором — хрящовим опорним органом[17]. Радула є унікальною для молюсків і не має відповідників у тварин інших груп. Окрім радули нерідко також бувають розвиненими хітинові щелепи[26].
Їжа, що потрапила до рота, приклеюється до в'язкої слини, яка завдяки руху війок спрямовується до шлунку[22]. На загостреному кінці шлунку на межі з кишкою розташований простиль — конусоподібне, загострене дозаду утворення, складене з різних осадових частинок. Слина рухом додаткових війок спрямовується на простиль так, щоб він слугував у ролі своєрідної бобіни. Поки ще слина не досягла простилю, кислотність шлунку робить слину менш клейкою, і харчові частинки відокремлюються від неї[22]. Далі харчові частинки сортуються іншою групою війок. Менші частинки, переважно мінерали, спрямовуються війками до простилю, так що врешті-решт вони виділяються назовні, а великі частинки, здебільшого власне їжа, відправляються на перетравлення у сліпий відросток кишки. Процес сортування не є добре злагодженим[22]
Час від часу молюски виділяють фрагменти простилю, щоб запобігти його надмірному розростанню. Задня кишка відкривається анусом у мантійну порожнину. Анус омивається потоками води, що йде від зябер.
У хижих м'якунів травна система влаштована простіше[22]. У водяних молюсків є спеціальний орган — сифон, що є частиною мантії. Через сифон відбувається протікання води (рідше повітря), яке використовується для однієї або кількох цілей: руху, живлення, дихання, розмноження.
У деяких м'якунів роду Solemya травна система атрофована аж до повної редукції; припускають, що вони засвоюють поживні речовини за допомогою хемосинтезуючих бактерій.
Кровоносна система
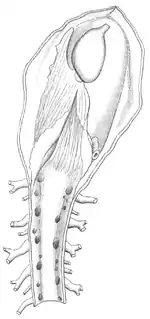
У молюсків незамкнена кровоносна система, до якої належать серце (орган, що забезпечує рух крові судинами та порожнинами тіла) та судини. Серце складається зі шлуночка та одного або, частіше, двох передсердь (у наутилусів чотири передсердя). Кровоносні судини виливають кров у проміжки між органами — у синуси та лакуни[26]. Потім кров знову збирається в судини й надходить до зябер або легень. Кров головоногих та деяких черевоногих молюсків на повітрі має незвичний блакитний колір. Цього кольору їй надає гемоціанін — дихальний пігмент, який містить мідь та виконує функції, подібні до функцій гемоглобіну в крові хордових та кільчастих червів, тому при окисленні кров стає блакитною[28].
У головоногих кровоносна система майже замкнена: поза судинами кров перебуває тільки коли частково з капілярів, вен та артерій витікає у дрібні лакуни[28].
Дихальна система
.jpg.webp)
Дихальна система представлена пірчастими шкірними адаптивними зябрами — ктенидіями. Також важливе значення має шкірне дихання, для деяких м'якунів воно навіть є єдиним. У наземних молюсків замість ктенидіїв є особливий орган повітряного дихання — легеня, що являє собою видозмінену мантійну порожнину, стінки якої пронизані кровоносними судинами[26][29].
Видільна система
Видільна система молюсків складається з нирок, які є видозміненими метанефридіями[25]. У них накопичуються продукти виділення у вигляді грудочок сечової кислоти, які виводяться один раз на 14–20 днів. У багатьох черевоногих є лише одна, ліва нирка, а найбільшу кількість нирок (5–6 пар) мають представники моноплакофор[30]. Внутрішнім кінцем (заглибиною) кожна нирка відкривається в навколосерцеву сумку (перикард), а зовнішнім (видільними отворами) — у мантійну порожнину, куди виводяться кінцеві продукти обміну речовин[26]. Передсердя молюсків, фільтруючи кров, фактично є частиною видільної системи.
Осморегуляція
Морські молюски є пойкілосмотичними тваринами, тобто вони не здатні зберігати постійний осмотичний тиск (ОТ) у тканинах у разі зміни солоності води[31], й ОТ крові в них змінюється зі зміною тиску в навколишньому середовищі, стаючи йому рівним (іншими словами, ОТ морських молюсків дорівнює ОТ морської води, тобто вони ізотонічні тому середовищу, у якому мешкають). Постійний вміст води та солей у клітині в них забезпечує клітинна осморегуляція: при збільшенні або зменшенні ОТ середовища на таку ж величину змінюється концентрація осмотично активних органічних речовин (переважно амінокислот). У такий спосіб ОТ у клітині та в зовнішньому середовищі зрівнюються[32].
Прісноводні молюски гіпертонічні до свого середовища мешкання, оскільки їхній ОТ більший за ОТ прісної води. У зв'язку з цим проблема осморегуляції постає більш гостро, ніж у морських молюсків. Загальною рисою для прісноводних молюсків є те, що вони мають рівень солоності тканин набагато нижчий, ніж у морських, а також інших прісноводних тварин; крім того, у прісноводних двостулкових молюсків цей показник є найнижчим серед усіх тварин[33]. Тож різниця між ОТ молюска та середовища не дуже велика, проте необхідність осморегуляції зберігається. Цю функцію виконують метанефридії, виділяючи разом із сечовою кислотою надлишки води та солей[34]
Нервова система
Нижчим групам молюсків (Caudofoveata, борозенчасточеревним та панцирним) притаманна нервова система драбинчастого типу, подібна до такої в деяких кільчастих червів[26]. Вона складається з навкологлоткового кільця та чотирьох стовбурів: двох педальних (іннервують ногу) та двох вісцеральних[35] (іннервують вісцеральну масу).
У більшості інших представників молюсків спостерігається утворення гангліїв та їх зміщення до переднього кінця тіла, причому найбільше розвивається надглотковий нервовий вузол («головний мозок»)[19]. У результаті формується нервова система розкидано-вузлового типу.
У нервовій системі розкидано-вузлового типу є дві (у двостулкових — три) пари нервових ланцюгів: двоє черевних ланцюгів іннервують внутрішні органи, а двоє педальних — ногу. Обидві пари ланцюгів містять ганглії як локальні центри контролю важливих частин тіла. Більшість пар відповідних гангліїв, розташованих по обидва боки тіла, з'єднана між собою комісурами. Зазвичай є 5 пар гангліїв: церебральні (іннервують очі та щупальця), педальні (ногу), плевральні (мантію), парієтальні (органи дихання та осфрадії) та вісцеральні (внутрішні органи). Іноді виділяють також букальні ганглії, що іннервують глотку. Вони винесені з навкологлоткового кільця й розташовуються на дорсальному боці глотки в місці переходу її в стравохід[36]. Церебральні, педальні та вісцеральні ганглії пов'язані між собою поперечними нервовими тяжами — комісурами[37]. Майже всі ганглії розташовуються нижче кишки, винятком є лише церебральні ганглії, розташовані над стравоходом. Педальні ганглії локалізовані відразу під стравоходом, а їхні комісури та конективи, що зв'язують їх із церебральними гангліями, утворюють нервове кільце навколо стравоходу. У видів, що мають мозок, він оточує стравохід кільцем[22].
У багатьох черевоногих через закручування тулуба утворюється перехрещення між плевральними та парієтальними гангліями. Це перехрещення має назву хіастоневрія. Нервову систему без перехрещення називають епіневральною, а з перехрещенням — хіастоневральною[38].
Окрім рефлекторної діяльності, нервова система також регулює ріст та розмноження за допомогою різних нейрогормонів[39].
Органи чуття
До органів чуття молюсків належать очі та щупальця, розташовані на голові, органи хімічного чуття — осфрадії, розташовані поряд з основою зябер, та статоцисти на нозі[25]. Акомодація ока (у видів, які до неї здатні) відбувається шляхом зміни його форми — віддалення або зближення сітківки та кришталика. Будова ока в головоногих дуже подібна до такої у хребетних тварин, проте його акомодація відбувається інакше і вони по-різному розвиваються у процесі онтогенезу. Переважно на голові, нозі та на краю мантії зосереджені дотикові сенсорні клітини[26].
Статева система

Усі легеневі молюски — гермафродити, більшість зябродихаючих — роздільностатеві. За гермафродитизму кожна особина виконує роль і самиці, й самця, при спаровуванні двох особин відбувається перехресне запліднення. Протоки гонад (гонодукти) є целомодуктами[26]. Ними статеві клітини прямують у целом, звідки вони відфільтровуються нирками і спрямовуються в мантійну порожнину. Такий механізм існує у роздільностатевих молюсків із зовнішнім заплідненням, яке відбувається у воді. У більш розвинених головоногих та більшості черевоногих м'якунів запліднення внутрішнє. Восьминоги для перенесення статевих продуктів у мантійну порожнину самиці використовують спеціалізоване видозмінене щупальце — гектокотиль[40].
Життєвий цикл
Після запліднення яйцеклітина (зигота) молюсків зазнає спірального дроблення, що є однією з ознак типу. У головоногих яйця проходять неповне дроблення[30]. Розвиток може бути прямим або непрямим. Деякі молюски є живородними, наприклад, черевоногі з родини Viviparidae. Яйця цих м'якунів запліднюються в тілі самиці, а потім розвиваються у яйцеводі; новонароджені равлики починають вести спосіб життя як дорослі особини[41].
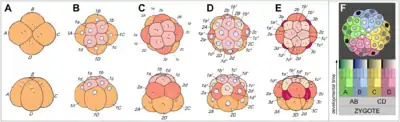
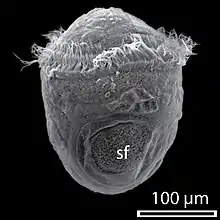
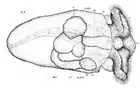
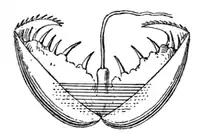
У нижчих молюсків з яйця виходить трохофорна личинка, у більшості ж ця стадія проходить у яйці[26]. Трохофора веде планктонний спосіб життя, живиться харчовими частинками що плавають у товщі води, спрямовуючи їх до рота за допомогою двох рядів війок, розміщених на «екваторі» її тіла. Крім цього, рух війок створює течію води для видалення з анального отвору неперетравлених решток їжі. Нові тканини утворюються зі смуг мезодерми, які розташовані всередині так, що апікальний пучок війок та анус віддаляються один від одного у процесі росту личинки.
Черевоногим, двостулковим та лопатоногим молюскам притаманна личинка, яка має назву вітрильник або велігер[35], оскільки має широкі лопаті, на яких розташовані війки, і нагадує вітрило. Ці лопаті розвинулися з екваторіальної смуги війок, щонайближчих до апікального пучка війок. Згодом личинка осідає на дно і зазнає метаморфозу, перетворюючись на дорослу особину.
Деякі прісноводні двостулкові мають личинку, що називається глохідій[42]. Глохідій має двостулкову мушлю з гачками на краях, потужний адуктор та клейку нитку. За допомогою нитки та гачків глохідії прикріплюються до зябер та шкіри риб, що пропливають повз, та занурюються в їхні тканини. Навколо глохідію утворюється невелика пухлина, у якій личинка живиться тканинами господаря та перетворюється на маленького молюска. Після закінчення метаморфозу маленькі молюски випадають назовні через розрив пухлини та осідають на дні водойми. Тимчасовий ектопаразитизм личинок двостулкових забезпечує їхнє розселення[43][44].
Більша частина прісноводних молюсків, усі наземні види, а також деякі морські, зокрема всі головоногі, розвиваються без метаморфозу, тобто в них відбувається прямий розвиток[26][30].
Деякі головоногі турбуються про потомство[45].

Тривалість життя молюсків дуже варіює. У той час як багато видів молюсків живуть кілька місяців або навіть тижнів[47], серед двостулкових є представники з найбільшою тривалістю життя серед усіх тварин — понад 400 років. До цього класу також належить більша частина видів тварин, чия тривалість життя зазвичай становить до 150 років. Так, у морського м'якуна Panopea abrupta, океанського Arctica islandica та прісноводної річкової скойки максимальна тривалість життя становить 163, 405 та 190 років відповідно[46].
Поширення
Молюски поширені на всій Земній кулі: заселяють суходіл, мілководдя та глибини морів та океанів[48], особливо численні у прибережній зоні[30]. Порівняно невелика кількість видів живе у прісних та солонуватих водоймах. Найбільше розмаїття м'якунів спостерігають у тропічних морях. Найглибоководніші молюски виявлені в Маріанській западині на глибині приблизно 11 000 м[49], а на суходолі равлики поширені від тундри до тропіків та від низовин до верхніх поясів гір[30].
Спосіб життя та поведінка
Молюски становлять численну й розмаїту групу тварин, спосіб життя яких також дуже різноманітний.
Хітони — рухливі, повільно плазуючі тварини; живляться водоростями, яких зішкрібають радулою з твердих поверхонь. Є хижі види[50]. Представники класу борозенчасточеревних утворюють високоспеціалізовану групу, вони мають червоподібне тіло й мешкають на гідроїдних та коралових поліпах[51]. Ямкохвості — морські червоподібні молюски, мешкають у ґрунті та живляться детритом або форамініферами[52]. Лопатоногі — вузькоспеціалізовані рийні організми[53]. Перші сучасні моноплакофори (Neopilina galatheae) були виявлені в Тихому океані на глибині 3590 м[54], тобто вони є морськими глибоководними організмами.
Черевоногі
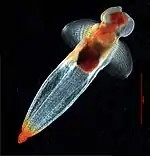
Черевоногі — найчисленніший та найрізноманітніший клас молюсків, вони опанували всі середовища існування, окрім повітряного. Черевоногі здебільшого є мешканцями морів, де поширені як у прибережній зоні, так і на великих глибинах, у найрізноманітніших температурних умовах[55]. Більшість з них повзає поверхнею дна, деякі ведуть рийний спосіб життя. Є черевоногі, які втратили мушлю й перейшли до плавучого способу життя: крилоногі (Pteropoda) та кіленогі (Heteropoda)[56]. Вони мають склоподібне та прозоре тіло, нога перетворилася на два великі крилоподібні плавці (у крилоногих) або утворює єдиний кілеподібний плавець (кіленогі)[55].
Серед них є наземні види, що дихають за допомогою легень. Багато легеневих переносять різкі коливання температури, впадаючи у сплячку — зимову на півночі, літню та зимову на півдні. Равлики переносять несприятливі умови у ґрунті, втягуючись у черепашку та заклеюючи устя застиглою слизовою плівкою з великим вмістом вапна — епіфрагмою[55]. У деяких легеневих (слизняків) мушля редукована. Слизняки можуть зариватися у ґрунт або прогризати порожнини у тканинах рослин, грибів та лишайників[57]. Є також хижі слизняки, що живляться іншими молюсками або олігохетами[58].
Серед прісноводних молюсків є також вторинноводні, які дихають повітрям, а живуть у воді (ставковики, котушки)[56].
Також є дуже невелика кількість паразитичних черевоногих, як, наприклад, Eulima bilineata та Odostomia turrita[59], Entocolax, Parenteroxenos. Справжні паразити є лише серед передньозябрових, які переважно паразитують на шкірі або в порожнині тіла голкошкірих. Паразитичні черевоногі через свій спосіб життя мають спрощену організацію: вони втратили мушлі, мантії, ноги та інші органи аж до повної редукції травної, кровоносної та нервової систем[55].
Особливу групу черевоногих становлять коралобіонти (Coralliophilidae); їхня подібна до звивистої трубки мушля приростає до скелетів коралових поліпів. Верметиди зі схожими трубкоподібними черепашками формують щільні поселення на твердому субстраті[60].
Морські блюдечка, що живуть у прибійній смузі, мають виражений інстинкт «дому» (хомінг): під час припливу вони повзають камінням, зішкрібаючи водорості, а за відпливу повертаються щоразу на одне й те ж місце[61].
Двостулкові

Двостулкові здебільшого біофільтратори[60], ведуть малорухливий або нерухомий спосіб життя. Деякі при цьому прикріплюються до скель або водоростей бісусом (Mytilus, Pinna тощо), інші ж міцно приростають до субстрату стулкою мушлі (Ostrea, Pinctada)[62]. Проте морські гребінці здатні до реактивного руху, ляскаючи стулками мушлі завдяки потужному м'язу-замикачу; так гребінці можуть перепливати на невеликі відстані[63][64].
Молюски-каменеточці з родини Морських фініків за допомогою спеціального кислого секрету мантійної залози проточують ходи у вапняку й закріплюються в них за допомогою бісусу, виставляючи назовні сифони[63]. М'якуни з родини Pholadidae також здатні пошкоджувати вапняки, пісковики та навіть бетон, а молюски-деревоточці (Teredinidae) пошкоджують занурені у воду дерев'яні частини човнів, кораблів та гідротехнічних споруд[65].
Представники надряду Septibranchia ведуть переважно хижий спосіб життя[65].
Двостулкові з роду Entovalva ведуть ендосімбіотичний спосіб життя і трапляються лише у стравоходах голотурій[66].
Головоногі
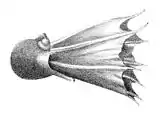
Головоногі — здебільшого хижі морські тварини, що вільно плавають у товщі води[67]. Багато головоногих здатні до реактивного руху. Деякі головоногі ведуть придонний спосіб життя, часто ховаючись у щілинах між камінням. Серед глибоководних форм відомі м'якуни зі схожими на телескопи очима, що стирчать[68].
Наутилуси є бенто-пелагічними молюсками, каракатиці ведуть некто-бентичний спосіб життя[69], а кальмари — головним чином нектонні тварини, що завдяки торпедоподібному тілу активно плавають у товщі води[70]. Більшість восьминогів веде придонний спосіб життя, але серед них є нектонні та навіть планктонні організми[70]. У деяких донних восьминогів між щупальцями розташована тонка перетинка, що надає тварині вигляд диску, що лежить на дні[68].
Живлення
За типом живлення молюсків поділяють на фільтраторів, рослиноїдних, хижих, паразитів та падальників, їхні раціони надзвичайно різноманітні.
Рослиноїдні молюски живляться водоростями. Одні з них живляться мікроскопічними нитчастими водоростями, яких вони зішкрібають за допомогою радули з різних поверхонь. Інші живляться макроскопічними рослинами, наприклад, бурими водоростями, зішкрібаючи радулою їхню поверхню. В іншому випадку більші рослини з'їдаються частіше, ніж дрібніші, оскільки рослина має бути великою настільки, щоб на ній умістився молюск[71].
Головоногі молюски здебільшого (а можливо, й повністю) є хижаками, тому в них радула має менше значення в порівнянні зі щупальцями та щелепами. Моноплакофори роду Neopilina використовують радулу звичайним чином, але до їхнього раціону входять найпростіші, наприклад, ксенофіофори Stannophyllum[72]. Голозяброві з групи Doridacea, а також деякі черевоногі з групи Vetigastropoda живляться губками[73][74]. Є молюски, які живляться гідроїдними поліпами[75]. Черевоногий хижий молюск Charonia tritonis живе серед коралових рифів і є одним із природних ворогів морської зірки «терновий вінець», яка руйнує корали[76].

Деякі молюски мешкають у симбіозі з бактеріями, які залучені у процес їхнього травлення, або ж живляться за їх рахунок. Наприклад, у деяких двостулкових м'якунів роду Solemya, що мешкають у ґрунті, де багато сірководню (узбережжя, зайняті звалищами, промисловими підприємствами) травна система атрофується різною мірою, аж до повної редукції. Живляться ж такі молюски, ймовірно, бактеріями, які окислюють сірководень, що оселяються в зябрах молюска або вільно живуть у ґрунті, але яких молюск відфільтровує й перетравлює у клітинах зябер[77]. Молюски-деревоточці (представники родини двостулкових молюсків Teredinidae) мають червоподібне тіло й живляться зануреною у воду деревиною. У цей субстрат вони вгризаються свердлувальним апаратом черепашки, яка в них редукована до двох маленьких пластинок на передньому кінці тіла. Через це деревина, пронизана численними ходами молюска-деревоточця, стає схожою на губку й легко руйнується. Перетравлювання деревини в молюсків-деревоточців здійснюють симбіотичні бактерії[78].
Є також «фотосинтезуючі» молюски — задньозяброві черевоногі з ряду Sacoglossa[79]. У їхніх тканинах накопичуються хлоропласти з'їдених водоростей, через що молюск набуває зеленого кольору й здатності до фотосинтезу. Це явище отримало назву клептопластія.
У деяких голозябрових черевоногих молюсків, що живляться гідроїдними поліпами, у порожнину спинних зябер вдаються зсередини відростки печінки, які містять неперетравлені жалкі клітини поліпів. У разі подразнення ці клітини вистрілюють і слугують для захисту молюсків[57].
Екологія
Значення в екосистемах
Через те, що молюски — надзвичайно численна та різноманітна група, представники якої пристосувалися до найрізноманітніших середовищ існування та ведуть найрізноманітніший спосіб життя, від фільтраторів до хижаків та паразитів, їхні ролі в екосистемах також широко варіюють.
Морські молюски складають значну частину бентосу. Щільність молюсків на морському дні може досягати значення декількох тисяч особин на 1 м². У товщі морської води численні активно плаваючі головоногі, що ведуть хижий спосіб життя[80].
Молюски складають важливу ланку в ланцюгах живлення у водних і наземних екосистемах; спектр їхнього живлення надзвичайно широкий[80]. Дуже незначна кількість видів пристосувалася до паразитичного способу життя.
Особливо велика роль двостулкових молюсків як біофільтраторів, що очищають водойми від органічного забруднення. Крім того, вони поглинають та накопичують у своєму тілі важкі метали. Велика роль двостулкових і в утворенні осадових порід[81].
Природні вороги
Природні вороги наземних молюсків (тобто легеневих равликів) надзвичайно різноманітні. Ними живляться птахи, плазуни, ссавці, а також жуки родин Carabidae, Staphylinidae, Lampyridae, Drilidae та Silphidae. Деякі двокрилі теж є хижаками та паразитоїдами наземних молюсків. Інші хижаки наземних равликів: деякі планарії, хижі наземні равлики, двопарноногі та губоногі багатоніжки, напівтвердокрилі та павуки. На наземних молюсках паразитують деякі кліщі, нематоди, інфузорії, мікроспоридії[82].

Для захисту від хижаків морські молюски нерідко виділяють хімічні речовини, що діють на органи хімічного відчуття хижака. Наприклад, у каракатиць та інших головоногих є спеціальні чорнильні мішки, щоправда, чорнило вони використовують тоді, коли інші засоби — відкидання щупальця та секреція отрути — не спрацьовують. За допомогою таких речовин, наприклад, захищаються від своїх потенційних ворогів (птахів, риб, ракоподібних, актиній) черевоногі молюски з роду Aplysia. Основними ворогами головоногих молюсків також є риби та ракоподібні[83].
Міцна округла мушля робить двостулкових молюсків малодоступними для хижаків. Проте багато тварин, тим не менш, живляться ними. Серед них риби, такі як короп[84], та птахи, наприклад, кулики-сороки та лелеки-роззяви, які розкушують мушлі пристосованим для цього дзьобом[85], та сріблястий мартин, що розбиває черепашки, кидаючи на них каміння[86]. Калани живляться багатьма двостулковими молюсками, розбиваючи їхні мушлі за допомогою каміння[87]. Одним з головних хижаків арктичних вод, що харчуються двостулковими молюсками, є морж[88]. З безхребетних природними ворогами двостулкових є краби[89], морські зірки та восьминоги[90]. Деякі черевоногі молюски — Urosalpinx, Eupleura caudata, Polynices heros, Neverita duplicata — також знищують двостулкових молюсків, зокрема цінних промислових тварин — устриць та мідій[91].
Механізми захисту від ворогів у двостулкових молюсків різноманітні. Деякі закопуються у ґрунт, як, наприклад, Siliqua patula, яка здатна закопатися за 7 секунд[92]. Морські гребінці та деякі інші молюски здатні плавати, ляскаючи стулками мушлі. Інші двостулкові молюски здатні відстрибувати від загрози на нозі, використовуючи її, як пружину[93]. Двостулкові, які мають сифони, можуть сховатися всередині мушлі, виставивши сифони назовні; якщо хижак обламає їх, вони регенеруються[94]. М'якуни з роду Limaria у разі роздратування виділяють токсичні речовини[95].

Симбіонти
Є випадки симбіозу між молюсками та іншими безхребетними. Наприклад, черевоногий молюск Colus gracilis входить у симбіоз з актинією Hormathia digitata, яка при цьому оселяється на мушлі молюска[96].
Описані також симбіотичні взаємовідносини між молюсками та найпростішими (одноклітинними водоростями). Зокрема, двостулковий молюск Corculum cardissa входить у симбіоз із найпростішими Symbiodinium corculorum з групи динофлагелят. Клітини найпростіших виявляють світловою або трансмісійною електроскопією переважно у тканинах мантії та зябер молюска. Найпростіші (зооксантели) поселяються також у потовщеному краї мантії велетенської тридакни, але, на відміну від Corculum cardissa, у клітинах зябер вони не виявлені[97][98].
На мушлях молюсків розвивається одна зі стадій життєвого циклу червоних водоростей з роду Porphyra[99].
Біолюмінесценція
Деякі морські молюски здатні до біолюмінесценції. До них належать кілька незвичайних черевоногих, наприклад, м'якуни роду Planaxis та ефектні голозяброві слимаки роду Phylliroe[100], а також двостулкові, такі як представники роду Pholas[101]. М'якун Hinea brasiliana використовує дифузну біолюмінесценцію аби в очах хижака здаватися більшим і в такий спосіб його відлякати[100].
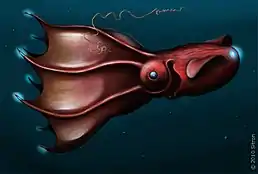
Проте найбільша кількість молюсків, здатних до біолюмінесценції, належить до класу Головоногих. Лише серед кальмарів їх налічують щонайменше 70 видів[102]. Декілька родів з родин Sepiolidae та Loliginidae випромінюють світло завдяки бактеріям-симбіонтам[103][104][105][106]. Решта кальмарів здатні світитися самостійно, використовуючи люмінесцентну речовину люциферин та фермент люциферазу, що каталізує окислення люциферину[101]. Кальмари демонструють велике розмаїття структур, що беруть участь у біолюмінесценції. Більшість має 2 вентральних фотофори — органи, що випромінюють світло[107]. Глибоководний молюск Vampyroteuthis має настільки своєрідні органи світіння, що його навіть виокремили в самостійний ряд. На додачу до двох великих мантійних фотофорів та маленьких світних органів, розкиданих усім тілом, він здатний випромінювати світло спеціальними органами на кінцях щупалець; ймовірно так він збиває з пантелику хижаків[108]. Кальмар Octopoteuthis має велику кольорову розмаїтість забарвлення, використовує кінці щупалець, які випромінюють світло, як принаду для здобичі. Якщо його потурбувати, він здатен відкидати щупальця[109]. Дуже активний молюск Taningia danae має клешнеподібні гачки на щупальцях замість присосків та великі (до 2 м) органи світіння на кінцях щупалець. Припускається, що він використовує люмінесценцію для внутрішньовидової комунікації, а також засліплення здобичі[101].
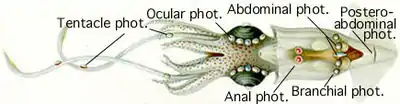
Восьминоги також здатні до біолюмінесценції. Самиці пелагічних глибоководних восьминогів родів Japetella та Eledonella мають зеленувато-жовте кільце навколо рота, яке тільки періодично люмінесціює[110]; це може відігравати роль у розмноженні[111]. У Stauroteuthis та інших родів глибоководних восьминогів є присоски, що випромінюють світло. Вважають, що сяючі лінії, розташовані вздовж щупалець, використовуються для принаджування здобичі[112].
Отже, розмаїття світних структур у головоногих, як і в молюсків загалом, надзвичайно велике, і кількість їх незалежних появ у процесі еволюції значно більша, ніж вважається[101].
Захворювання молюсків
Молюски схильні до низки вірусних, бактеріальних та паразитичних захворювань. Прикладами вірусних захворювань молюсків можуть бути вірусний гангліоневрит абалонів (англ. abalone viral ganglioneuritis (AVG))[113] та герпесоподібна вірусна інфекція (англ. herpes-like virus infection); бактеріальних — хвороба коричневого кільця (англ. brown ring disease) та фіброз личинок та молодих особин (англ. larval/juvenile fibrosis); паразитичних — перкінсоз (англ. perkinsosis), мартеіліоз (англ. marteiliosis), бонаміоз (англ. bonamiosis), гаплоспоридіоз (англ. haplosporidiosis) та митіліколоз (англ. mytilicolosis)[114].
Механізми захисту в молюсків наразі значною мірою залишаються нез'ясованими. Наприкінці XX століття вважалося, що в молюсків відсутні імуноглобуліни (антитіла) та набутий імунітет. Головним механізмом захисту вважався фагоцитоз. Проте останнім часом встановлено велику різноманітність типів кров'яних клітин (гемоцитів) молюсків та їхні відмінності в різних груп м'якунів; так, в абалонів та морських гребінців не були виявлені гранулоцити, а в інших черевоногих — виявлені. Походження, життєвий цикл, період життя, функції кожного типу клітин ще належить визначити[115].
Деякі збудники захворювань молюсків здатні впливати на імунну відповідь господаря, модифікуючи її. Наприклад, мікроклітини Bonamia roughleyi стимулюють фагоцитоз себе гемоцитами господаря, що підходять. Проте бактерію це не вбиває, навпаки, її клітини продовжують розмножуватися усередині клітини-господаря, зрештою лізуючи її оболонку та вивільняючись назовні. Це призводить до масового руйнування кров'яних клітин та загибелі господаря — устриці[115].
Відомі випадки, коли хімічні речовини, що виділяються патогеном, каструють молюска. Наприклад, речовини, що виділяються спороцитами Zoogonius rubellus, каструють його господаря — морського черевоногого Nassarius obsoletus. Дослідження прісноводних равликів Lymnaea stagnalis, заражених трематодою Trichobilharzia ocellata, показали, що речовини, які секретуються трематодою, змінюють експресію генів господаря, що пригнічує мітотичні поділи в чоловічому копулятивному органі та стимулює розвиток жіночих спинних ендокринних тілець. Схожа ситуація спостерігається в молюска Haliotis asinina та трематоди Allopodocotyle. Шистосома Schistosoma mansoni використовує для власних потреб нейромедіатори серотонін та дофамін господаря, Biomphalaria glabra, викликаючи, зміни в його ендокринній системі[115].
Систематика
Тип Молюски (Mollusca) деякі автори поділяють на 2 систематичні групи, яким іноді надають ранг підтипу: Боконервові (Aculifera), що об'єднує панцирних та безпанцирних молюсків, та Черепашкові (Conchifera), яка включає всі інші сучасні класи[116][50][117].
Серед науковців-малакологів є різні думки щодо кількості класів молюсків. Найчастіше описують 8 сучасних[118] та принаймні 2 викопних[119] класи м'якунів:




- Ямкохвості (Caudofoveata) — дрібні червоподібні тварини, які мешкають у товщі пухкого морського осаду на глибинах від 200 до 3000 м[120], де займають екологічну нішу вибіркових детритофагів або хижаків. Довжина тіла зазвичай становить 1—15 мм, деякі особини можуть досягати 30 мм[121]. Клас нараховує 120 сучасних видів[118].
- Борозенчасточеревні (Solenogastres) — морські молюски, які мешкають переважно в колоніях гідроїдних та коралових поліпів, по яких повзають, згинаючи червоподібне тіло. Трапляються на глибинах від 200 до 3000 м[120]. У зв'язку з вузькою спеціалізацією до перебування на колоніях поліпів у більшості редукована мантійна порожнина, зябра та частково нога; розвинений захисний кутикулярний покрив, шкірно-м'язовий мішок, спеціалізована радула[51]. Довжина тіла зазвичай становить від 0,5 до 25 мм[122], лише один вид сягає 25 см[123]. Нараховують 200 сучасних видів[118].
- Панцирні (Polyplacophora) — морські повільно плазуючі тварини; живляться, зішкрібаючи водорості з твердого субстрату, до якого присмоктуються підошвою ноги. Невелика кількість є хижаками. На спинному боці — панцир з 8 пластинок; у разі небезпеки можуть згортатися в кулю, подібно до їжака, виставляючи пластинки назовні. Мантійна порожнина має багато пар зябер. Нога оснащена підошвою[50][124]. Розміри від менше ніж 1 см[125] до 36 см за ваги до 2 кг (Cryptochiton stelleri)[126]. Клас нараховує приблизно 1000 сучасних видів[118].
- Моноплакофори (Monoplacophora) — один із найпримітивніших класів м'якунів, представники якого мають архаїчні риси — збережені великі целомічні порожнини, метамерію в будові деяких систем органів, примітивну нервову систему[127]. Мушля має вигляд шапочки. Розміри дрібні, лише кілька сантиметрів завдовжки. Глибоководні морські молюски, мешкають на глибинах від 1800 до 7000 м, один вид трапляється на глибині приблизно 200 м[120]. Відомий 31 сучасний вид[118].
- Двостулкові (Bivalvia) — виключно водяні, прісноводні та морські м'якуни, найхарактерніші особливості будови яких — черепашка, складена з двох стулок, розташованих з боків тіла, та редукція голови і всіх пов'язаних із нею утворень, включно з радулою. Складки мантії переважної більшості двостулкових молюсків утворюють на задньому кінці тіла ввідний та вивідний сифони, за допомогою яких двостулкові створюють потік води в мантійній порожнині[128][129]. Розміри стулок черепашки велетенської тридакни, найбільшого двостулкового молюска, можуть сягати майже 140 см, а вага до 333 кг[130]. Клас нараховує приблизно 20 000 сучасних видів[118].
- Лопатоногі (Scaphopoda) — виключно морські бентосні молюски, що мешкають від літоралі до глибин у 7000 м[131]. Довжина тіла від 1,5 мм до 15 см. Тіло двосторонньо-симетричне, поміщене у трубчасту мушлю, що нагадує вигнуте ікло або бивень слона. Нога (у деяких видів редукована) зазвичай оснащена придатками у вигляді пари бічних лопатей або зубчастого диска та пристосована до риття у ґрунті. На голові є рот та численні ниткоподібні придатки (каптакули), призначені для пошуку та захоплення їжі. Глотка зі щелепою та радулою (по 5 зубів у кожному сегменті). Зябра скорочені, очей немає. Кровоносна система лакунарного типу, циркуляція крові відбувається завдяки скороченням ноги[132]. Нараховують приблизно 500 сучасних видів[118].
- Черевоногі (Gastropoda) — найчисленніший клас молюсків, до якого належать приблизно 70 000 (80 % усіх сучасних видів м'якунів)[118]. Головна ознака представників класу — торсія, тобто поворот вісцерального мішка на 180 °. Крім того, гастроподам здебільшого притаманна наявність турбоспіральної черепашки. Черевоногі м'якуни широко розповсюджені на суходолі, у прісних та морських водоймах[133]. Розміри черевоногих складають від 0,5 мм (деякі наземні равлики, найменші серед молюсків) до 90 см (довжина мушлі) за ваги до 18 кг (Syrinx aruanus)[134].
- Головоногі (Cephalopoda) — виключно морські молюски, яким властиві двостороння симетрія та 8, 10 або більша кількість щупалець навколо голови, що розвинулися з ноги[135]. У представників підкласу Coleoidea, або двозябрових, мушля редукована чи повністю відсутня, тоді як у представників Nautiloidea збереглася зовнішня мушля. Головоногі мають найбільш досконалу з безхребетних кровоносну та найбільш розвинену нервову системи[136]. До головоногих належать наутилуси, кальмари, каракатиці та восьминоги. Розміри головоногих молюсків складають від 1 см довжини за ваги приблизно 0,2 г у каракатиць роду Idiosepius[137] до 10 м довжини й ваги до 495 кг у велетенських кальмарів[7], найбільших безхребетних тварин. Описано приблизно 900 сучасних та 11 000 викопних видів[118].
- † Rostroconchia — викопні морські молюски. Можливі предки двостулкових[138].
- † Helcionelloida — викопні морські молюски схожі на равликів, на кшталт Latouchella[139].
Порівняльна характеристика класів молюсків
У нижче наведеній таблиці представлено порівняльну характеристику будови представників сучасних класів молюсків (класи Caudofoveata та Solenogastres у даній таблиці об'єднані в один клас — Aplacophora):
| Класи | |||||||
| Параметр порівняння[140] | Безпанцирні (Aplacophora)[120] | Панцирні (Polyplacophora)[124] | Моноплакофори (Monoplacophora)[141] | Черевоногі (Gastropoda)[14] | Головоногі (Cephalopoda)[142] | Двостулкові (Bivalvia)[143] | Лопатоногі (Scaphopoda)[131] |
|---|---|---|---|---|---|---|---|
| Радула (тертушка) | Відсутня у 20 % Neomeniomorpha | + | + | + | + | Немає | Внутрішня, не може висуватися за межі тіла |
| Широка м'язиста нога | Редукована або відсутня | + | + | + | Перетворена на щупальця | + | Маленька, лише на «передньому» кінці |
| Нутрощевий мішок (вісцеральна маса) | Не виражений | + | + | + | + | + | + |
| Великий сліпий відросток кишки | У деяких відсутній | + | + | + | + | + | Немає |
| Великі метанефридії (нирки) | Немає | + | + | + | + | + | Невеликі, прості |
Еволюція
Викопні
Викопні знахідки свідчать про появу в кембрійському періоді палеозойської ери черевоногих, двостулкових та головоногих молюсків. Деталі походження молюсків як типу від корінної групи Lophotrochozoa та їхньої подальшої диверсифікації до відомих нам сучасних та вимерлих класів наразі остаточно не з'ясовані та є предметом суперечок вчених.
Одним з головних спірних питань є систематичне положення деяких представників викопної едіакарської та ранньокембрійської фауни. Наприклад, Kimberella (організм з відкладень віком 555 млн років) була описана Федонкіним як «молюскоподібна» тварина[144][145], проте інші дослідники охарактеризували її лише як «можливо двосторонньо-симетричну»[146][147].


Ще більш гостро стоїть питання, чи була молюском Wiwaxia (що жила 505 млн років тому), а точніше, чи був її ротовий апарат різновидом радули або ж він був ближче до ротового апарату поліхет[146][148]. Н. Баттерфілд, що не відносить Wiwaxia до молюсків, у той же час інтерпретував як фрагменти радули давніші (510—515 млн років) скам'янілості[149].
Схожі сумніви відсутні щодо належності до типу молюсків представників ряду Helcionelliformes, знайдені у відкладеннях віком 540 млн років у Сибіру та Китаї[151][152]. Їхні мушлі нагадують черепашки равликів. З цих знахідок можна зробити висновок, що молюски з мушлями з'явилися навіть раніше за трилобітів[150]. Більшість знайдених хелціонелід має довжину лише кілька міліметрів, але трапляються й екземпляри довжиною у кілька сантиметрів. Було припущено, що дрібні хелціонелліди — це молодняк, а великі — дорослі особини[153].
Деякі дослідники відносять хелціонеллід до перших черевоногих[154], але інші мають сумнів щодо цього через відсутність у них явних ознак торсії[155][156][157].
Тривалий час найбільш раннім головоногим вважали Volborthella віком понад 530 млн років, але виявлення особин, які збереглися краще, засвідчило, що цей вид, на відміну від молюсків, не секретував мушлю, а збирав її з крупинок двоокису кремнію. Крім того, його черепашки, на відміну від мушель головоногих, не розділені на камери. Таким чином, нині класифікація Volborthella залишається невизначеною[158].
Найдавнішим головоногим нині вважають пізньокембрійського Plectronoceras. Його мушля поділена на камери перетинками — септами й має сифон, як у сучасних наутилусів. Через наявність у мушлях Plectronoceras «баласту» (кам'янистих відкладень) дослідники зробили висновок, що цей молюск був придонним, а не вільно плаваючим, як сучасні головоногі[159]. Усі головоногі, які мають зовнішню мушлю (окрім наутилусів), вимерли до кінця мезозойської ери — 65 млн років тому[160].
Ранньокембрійських викопних Fordilla та Pojetaia вважають ранніми двостулковими[161][162][163][164]. Двостулкові молюски, що нагадують сучасних, з'являються в ордовицьких відкладеннях (443—488 млн років тому)[165].
У класифікації викопних молюсків осторонь стоїть клас Hyolitha. Маючи мушлі та навіть оперкулум, вони досить схожі на молюсків. Але деякі дослідники відносять цей клас до самостійного типу, хоча й не вказують точне положення цього типу на «дереві життя»[166].
Збільшення кількості екологічних ніш, освоєних молюсками, відбувалося поступово. У кембрійському періоді молюсків знаходять лише у морських відкладеннях. Їхнє розповсюдження у прісноводні водойми відбулося у девонському періоді, а перші суходільні молюски (легеневі равлики) виявлені лише в шарах, датованих кам'яновугільним періодом[167].
Філогенія
| Lophotrochozoa |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Філогенія (еволюційне дерево) молюсків також поки остаточно не визначена. На додачу до суперечок про те, ким є Kimberella та всі Halwaxiida — молюсками чи їхніми близькими родичами[145][146][148][149], суперечки виникають і з приводу родинних зв'язків між різними групами сучасних молюсків[147], оскільки деякі групи тварин, традиційно представлені у складі молюсків, можуть бути перевизначені як відмінні від молюсків, хоча й споріднені організми[169].
Молюски належать до групи Lophotrochozoa[147], яка характеризується наявністю трохофорної личинки й, у випадку лофофорат (Lophophorata), наявністю спеціального органу живлення — лофофору. Іншими представниками Lophotrochozoa є кільчасті черви (Annelida) та ще 7 типів морських тварин[170]. У кладограмі праворуч узагальнені дані з філогенії молюсків станом на 2007 рік.
Оскільки родинні зв'язки між членами цього еволюційного дерева нез'ясовані, складно виділити риси, успадковані всіма молюсками від останнього їхнього спільного предка[171]. Наприклад, незрозуміло, чи мав він метамерію (тобто чи складалося його тіло з однакових частин, які повторюються). Якщо це справді було так, то молюски походять від тварин, подібних кільчастим червам[172]. Думки вчених з цього питання розходяться: одні дослідники у 2006 році повідомили, що спостережувана метамерія в будові зябер та ретракторних м'язів ноги виникли вторинно[16], у той час як інші у 2007 році дійшли висновку, що предковий молюск мав метамерію, мав ногу для плазування та мінералізовану мушлю[147]. Вважається також, що мушля черепашкових молюсків пішла від спікул (невеликих голок) безпанцирних молюсків, проте це погано узгоджується з даними щодо онтогенезу спікул[171].
Мушля молюсків пішла від слизових покривів, які, поступово зміцнюючись, перетворилися на кутикулу. Кутикула непроникна для води та газів, тому з її появою став неможливим газообмін через покриви, що спричинило розвиток складнішого дихального апарату — зябер[150]. Надалі кутикула ставала більш мінералізованою, при цьому активізувався той самий транскрипційний фактор, що й у інших двосторонньо-симетричних тварин при формуванні скелету[172]. Перша мушля молюсків була укріплена майже виключно мінералом арагонітом[173].
Як зазначалося вище, родинні зв'язки між групами молюсків також є предметом обговорень. Кладограми, наведені нижче, зображають дві найбільш розповсюджені точки зору з цього питання.
Охорона молюсків

У Червоному списку Міжнародного союзу охорони природи (МСОП) станом на вересень 2013 року наведені дані про 6730 видів молюсків, з яких 310 вважаються вимерлими та ще 1885 видів (належать переважно до класу черевоногих) віднесені до категорій підвищеного ризику — EXW, CR, EN, VU[175]. Згідно окремого підрахунку для молюсків 2017 року, імовірно вимерлими з 1500 року є 1032 види цієї групи (більше половини всіх відомих сучасних вимирань видів), з них 803 — наземні молюски, 177 — прісноводні черевоногі, 46 — прісноводні двостулкові, 6 — морські черевоногі[8]. До Червоної книги України занесено 20 видів молюсків: 17 з класу черевоногих та 3 з класу двостулкових[176]. Переважно, види, які перебувають під загрозою зникнення різного ступеня ризику, є наземними та прісноводними[177].
Найбільшими небезпеками для водних м'якунів МСОП вважає забруднення навколишнього середовища (переважно рідкими відходами сільського господарства та деревообробної промисловості) та модифікацію природних водойм під потреби людини (будівництво дамб, відведення води)[175]. Ці та інші антропогенні чинники, у багато разів пришвидшуючи вимирання молюсків, здійснюють значно більший вплив на чисельність цих тварин, аніж природні. Особливо вразливі молюски-ендеміки островів, на кшталт представників ендемічної для Галапагоських островів родини черевоногих Bulimulidae, підродини Achatinellinae, ендеміків Гавайських островів тощо[177]. Так, на Гавайських островах до появи на них людини частота вимирання дорівнювала орієнтовно 1 виду на мільйон років. З появою на них полінезійців ця частота зросла до 1 виду на 100 років. З 1778 року, коли капітан Джеймс Кук відкрив Гаваї для європейців, частота вимирання досягла 1—3 видів на рік. На острові Муреа у Французькій Полінезії 8 видів та підвидів равликів роду Partula зникли менш ніж за 10 років через інтродукцію хижого равлика Euglandina rosea[177].
Зникаючим видам молюсків притаманні певні спільні риси: пізнє статеве дозрівання, порівняно велика тривалість життя, низька плодючість, обмежений ареал та специфічне середовище існування[177].
Проте деякі м'якуни завдяки людині значно розширили межі свого природного розповсюдження: велетенський африканський равлик Achatina fulica, уже згадуваний хижий Euglandina rosea, прісноводний двостулковий молюск Corbicula fluminea. Маючи надзвичайну плодючість та здатність пристосовуватися до різних умов, вони швидко захоплюють вільні екологічні ніші й наразі трапляються в найрізноманітніших місцях існування[177].
Значення для людини
Упродовж тисячоліть люди вживали молюсків у їжу. Окрім того, вони були джерелом різних цінних матеріалів: перлів, перламутру, пурпуру, тхелету та висону. У деяких культурах мушлі м'якунів слугували за валюту. Велетенські розміри та чудернацькі обриси деяких молюсків спричинили появу розповідей про морських чудовиськ, таких як кракен. Молюски певних видів отруйні й можуть становити небезпеку для людини. Є серед м'якунів і сільськогосподарські шкідники, наприклад, Achatina fulica.
Використання

- У харчовій промисловості
Молюски, особливо двостулкові, наприклад, мідії та устриці, здавна були для людини харчами[178]. Інші молюски, яких часто вживають у їжу, — восьминоги, кальмари, каракатиці та равлики[179]. У 2010 році в господарствах, що займаються аквакультурою, вирощено 14,2 млн т молюсків, що становить 23,6 % від усієї маси м'якунів, що були вжиті в їжу[180]. Деякі країни регулюють імпорт молюсків та інших морепродуктів, переважно для мінімізації ризику отруєння токсинами, що накопичуються в цих організмах[181].
За обсягами вилову черевоногі молюски поступаються двостулковим. У їжу вживають таких морських черевоногих, як морські блюдечка (Patella), абалони (Haliotis), сурмачі (Buccinum) (на Далекому Сході Росії здійснюють промисел, з них виготовляють консерви), літорини (Littorina), морські зайці (Aplysia). З наземних м'якунів у деяких країнах споживають равликів з родів Achatina, Helix, слизняків[182]. У деяких європейських країнах виноградних равликів розводять у спеціалізованих господарствах[55].
Обсяг вилову двостулкових молюсків на даний час поступається їхньому штучному розведенню в марикультурі. На спеціальних господарствах вирощують мідій та устриць, особливих успіхів такі господарства досягли в США, Японії, Франції, Іспанії, Італії. У Японії розвинена марикультура морської перлівниці з роду Pinctada[183]. Для місцевого населення країн карибського басейну, включно з Кубою, цінним промисловим молюском є велетенський стромбус (Lobatus gigas)[184].
Головоногі молюски також є об'єктами промислу, у їжу використовують м'ясо каракатиць, кальмарів та восьминогів. Каракатиць та деяких восьминогів виловлюють для отримання чорнильної рідини, з якої виготовляють натуральні туш і чорнило[185].
- У виробництві предметів розкоші та прикрас

.jpg.webp)
Більшість молюсків, які мають мушлі, утворює перли, проте комерційну цінність мають лише перлини, вкриті шаром перламутру. Їх утворюють лише двостулкові та деякі черевоногі молюски[143][14]. Серед натуральних перлів найбільшу коштовність мають перлини двостулкових молюсків Pinctada margaritifera та Pinctada mertensi, мешканців тропічної та субтропічної частин Тихого океану. Промисловий видобуток перлів у спеціалізованих господарствах полягає в контрольованому введенні твердих частинок в устриць і наступному збиранні перлин. Матеріалом для введення частинок часто слугують подрібнені черепашки інших молюсків. Використання цього матеріалу в промислових обсягах призвело до того, що деякі прісноводні види двостулкових молюсків у південно-східній частині США опинилися на межі вимирання[143]. Промислове розведення перлів стало поштовхом до інтенсивного вивчення хвороб молюсків, необхідного для забезпечення здоров'я поголів'я культивованих видів[186].


Перламутр, видобутий з мушель м'якунів, використовують для виготовлення різних виробів, наприклад, ґудзиків, а також для інкрустацій[187]. З гіпобранхіальних залоз деяких мурексів (Muricidae) видобувають пурпур, який за свідченням історика IV століття до н. е. Феофомпа, цінувався на вагу срібла[188]. Тхелет — барвник тваринного походження, яким у давнину фарбували тканини в синій, блакитний та пурпурно-блакитний колір. Ця речовина важлива для деяких обрядів юдаїзму як обов'язковий атрибут таких предметів як цицит (пасма бачення) та одяг первосвященика. Попри те, що метод отримання тхелету був втрачений у VI столітті н. е., на теперішній час у науковому світі склався консенсус, згідно з яким джерелом цієї речовини також був представник родини мурексів — Hexaplex trunculus[189]. Висон — коштовна тканина, сировиною для виготовлення якої є бісус — білковий матеріал, який виділяють деякі види двостулкових молюсків (найвідоміший — Pinna nobilis) для прикріплення до морського дна[190]. Прокопій Кесарійський, описуючи перські війни середини VI століття н. е., стверджував, що тільки представникам панівних класів дозволялося носити хламіди з висону[191].

Мушлі молюсків або окремі їхні частини використовували в деяких культурах як валюту. Вартість черепашок не була сталою, а залежала від їхньої кількості на ринку. Тому вони піддавалися непередбачуваним стрибкам інфляції, що були пов'язані зі знаходженням «золотої жили» або удосконаленням методів перевезення[192]. У деяких культурах прикраси з мушель були ознаками соціального становища[193].
- У косметології
У деяких косметичних салонах пропонують масаж з допомогою равликів. Спеціально вирощених равликів промивають у проточній воді кімнатної температури й висаджують на шкіру обличчя. Равлик залишає слиз та згрібає ороговілі лусочки шкіри з обличчя. Після прогулянки равлика по обличчю слиз розтирають, не змиваючи. Равликотерапія показана при неглибоких зморшках, мікропошкодженнями шкіри, рубцях і розтяжках[194].
- Як домашні тварини
У домашніх умовах найчастіше утримують наземних велетенських ахатин[195] та виноградних равликів. В акваріумістиці поширені ампулярії, меланії[196], котушки та ставковики[197]. У великих океанаріумах можна зустріти восьминогів, кальмарів та каракатиць[198].
- У науці
Високою специфічністю створюваного ними ефекту відрізняються токсини морських молюсків конусів (Conidae). Порівняно невеликі розміри їхніх молекул полегшують їх лабораторний синтез. Ці дві якості роблять токсини конусів об'єктом для досліджень в області неврології[199][200]. Молюски також становлять значний інтерес для розробки лікарських препаратів. Особливу увагу звертають до себе молюски, у травному тракті яких мешкають симбіотичні бактерії. Можливо, речовини, що виділяються цими бактеріями, знайдуть своє застосування як антибіотики або нейрологічні засоби[201].
Молюски є популярними лабораторними тваринами в нейробіології. Так, відкриття калієвих та натрієвих іонних каналів пов'язані з дослідженнями гігантського відростка нервової клітини (аксона) кальмара[202], їх подальші дослідження проводилися на нейронах черевоногих молюсків[203], розуміння молекулярних механізмів пам'яті поліпшилося після вивчення нервової системи аплізії.[204]
Мінералізовані мушлі молюсків добре зберігаються у викопному стані. Тому в палеонтології вони слугують «геологічними годинниками», керівними копалинами, які дозволяють з великою точністю проводити стратиграфічне датування шарів землі[205]
- Інше використання
Мушлі молюсків з давніх часів використовували як матеріал для виготовлення різних інструментів: рибальських гачків, різців, шкребків, насадок для мотики. Самі мушлі використовувалися як посуд, музичні інструменти (конх) та прикраси[187].
Черепашки переважно черевоногих, а також двостулкових та головоногих, є об'єктами поширеного у світі виду колекціонування, яке було започатковане ще в часи античності, а найбільшої популярності досягло в епоху Великих географічних відкриттів. У середині XIX століття в інтер'єрі вікторіанських будинків неодмінно була присутня засклена шафа, у якій поруч зі скам'янілостями та мінералами виставляли черепашки морських молюсків. Такий вид колекціонування залишається популярним і в наші дні[206].
Шкідники
Деякі види молюсків (переважно слизняки) є шкідниками сільськогосподарських культур[207]. Такий вид, потрапивши в нове середовище існування, здатний вивести з рівноваги місцеву екосистему. Наприклад, велетенська ахатина (Achatina fulica) — шкідник рослин, була інтродукована в багато областей Азії, а також на багато островів Індійського та Тихого океанів. У 1990-х роках цей вид досяг Вест-Індії. Спроба боротися з ним шляхом інтродукції хижого равлика Euglandina rosea тільки погіршила ситуацію: цей хижак ігнорує ахатин і замість них винищує місцеві види равликів[208].


Виноградний равлик шкодить винограду, а слизняки — городнім культурам[182]. Польовий слизняк (Agriolimax agrestis) завдає шкоди озимим посівам, картоплі, тютюну, конюшині, городнім рослинам, а Deroceras reticulatum завдає шкоди врожаям помідорів та капусти[58]. У південних районах загрозу для садів та городів становлять слизняки роду Parmacella[55].
Новозеландський прісноводний равлик Potamopyrgus antipodarum вперше був зареєстрований у Північній Америці в середині 1980-х років — спочатку в західних, а потім і в східних штатах США[209]. Хоча довжина цього равлика становить у середньому 5 мм[210], її виняткова плодючість призводить до концентрації до півмільйона особин на один квадратний метр, що зумовлює швидке вимирання місцевих молюсків та комах, а також риб, пов'язаних з ними харчовим ланцюжком[211].
Деякі молюски є ворогами промислових м'якунів. Наприклад, хижий равлик Crepidula fornicata іноді з'являється на устричних банках (тобто мілинах Північного моря та Атлантичного океану[62]) у таких кількостях, що самих устриць стає не видно; у результаті устриці гинуть[91].
Тередові, або молюски-деревоточці (Teredinidae) з класу двостулкових поселяються в зануреній у воду деревині, зокрема в підводних частинах дерев'яних човнів та кораблів, а також у стаціонарних гідротехнічних спорудах. У процесі життєдіяльності (див. вище розділ живлення), корабельний черв'як проточує в деревині численні ходи, сприяючи її швидкому руйнуванню[30]. Розмір збитку, який щорічно спричинюють корабельні черви, обчислюється мільйонами[212].
Річкова дрейсена (Dreissena polymorpha) часто оселяється в трубах та водогонах, прикріплюючись до твердого субстрату бісусом та утворюючи значні скупчення, чим призводить до їхнього засмічення[213].
Молюски та здоров'я людини

Багато молюсків самостійно виробляють або акумулюють з навколишнього середовища токсини, які становлять загрозу для здоров'я, а іноді й життя людини. Отруєння може відбуватися при укусі молюска, дотику до нього, або при вживанні його в їжу. Щоб зменшити цю загрозу, багато країн обмежують імпорт молюсків. Серед смертельно небезпечних молюсків можна зазначити деякі види конусів (Conidae) з класу Черевоногих та блакитнокільчастого восьминога (Hapalochlaena), який, щоправда, нападає на людину тільки якщо його спровокувати[214]. Тією чи іншою мірою отруйними є всі восьминоги[215]. Укус тропічного виду восьминогів Octopus apollyon викликає серйозне запалення, яке може тривати понад місяць навіть за правильного лікування[216]. Укус Octopus rubescens за неправильного лікування може викликати некроз тканин, а за правильного — справа може обмежиться тижнем головних болів та загальної слабкості[217].

Усі види м'якунів родини конусів (Conidae) є отруйними й можуть вжалити при дотику, проте представники більшості видів занадто малі, щоб становити серйозну небезпеку для людини[199]. Є задокументовані підтвердження численних випадків отруєнь, у тому числі зі смертельними наслідками[218]. Напевно, серйозну небезпеку для людини становлять лише деякі великі види, які здатні зловити й убити рибу[219].
Однак слід зазначити, що кількість людей, які гинуть від контакту з молюсками, становить менше ніж 10 % від кількості людей, що гинуть від контакту з медузами[218].
Є також двостулкові молюски, отруйні для людини; отруєння може супроводжуватися паралічем (англ. paralytic shellfish poisoning (PSP)), втратою пам'яті (англ. amnesic shellfish poisoning (ASP)), гастроентеритами, довготривалими неврологічними розладами та навіть смертю. Отруйність двостулкових зумовлена накопиченням ними одноклітинних, які виділяють токсини: діатомових водоростей або динофлагелят, яких вони відфільтровують з води; іноді токсини зберігаються навіть у правильно приготованих молюсках[220].
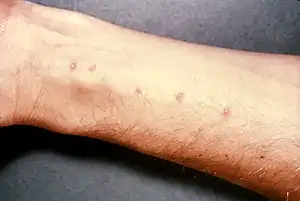
У тропічних країнах обох півкуль прісноводні равлики (зокрема родів Biomphalaria та Bulinus) є проміжними хазяями кров'яних сисунів (трематод) з роду Schistosoma, які викликають тяжке захворювання шистосомоз[221]. Малий ставковик, широко розповсюджений у Північній Євразії, є проміжним хазяєм печінкового сисуна, який паразитує у протоках печінки копитних ссавців та людини[222].
Образ молюсків у культурі
У культурах різних народів сформувалися певні стійкі стереотипи щодо представників трьох найпоширеніших та найвідоміших класів молюсків — черевоногих, двостулкових та головоногих.
Головоногі


Образ головоногого морського чудовиська є одним з найпопулярніших стереотипів молюсків. Міфічна істота кракен описувалася як велетенське головоноге створіння[223] настільки переконливо, що навіть увійшла до першого видання «Системи природи» Ліннея. Образ велетенського восьминога чи спрута, який становить небезпеку для кораблів та людей, був неодноразово зафіксований як у літературі (наприклад, сцена боротьби зі спрутами в романі Жуля Верна «Двадцять тисяч льє під водою»), так і в живописі та кінематографі (наприклад, у серії фільмів «Пірати Карибського моря»). Слово «спрут» стало стійким евфемізмом для позначення злочинної організації, як, наприклад, в однойменному телесеріалі. Сучасний образ головоногого чудовиська, Катацуморідако, був створений братами Стругацькими й описаний у романі «Хвилі гасять вітер»[224].
Черевоногі
Стереотип равлика в культурі — це, зазвичай, образ маленької, повільної та беззахисної істоти[225]. Образ «равлика на схилі», від якого нічого не залежить і який нічого не може змінити, використаний у назві однойменного роману Стругацьких, як алюзію на найвідоміший хайку Ісси («Тихо, тихо повзи, равлику, схилом Фудзі, вгору, до самої вершини!»). Разом з тим у Японії в образі равлика шанувався дух — «Господар води». Окрім того, равлики є центральними персонажами алтайської, бурятської, меланезійської міфології, а також міфології мешканців Науру[226]. У християнстві повільність равликів спричинила те, що вони стали символом одного зі смертних гріхів — ліні[227]. Мушля равлика в оповіданні Вірджинії Вульф «Kew Gardens» символізує розвиток сюжету за спіраллю[228].
Двостулкові

У давньогрецькій культурі двостулкові молюски були пов'язані з культом богині вроди й кохання — Афродіти. На картині італійського митця Боттічеллі «Народження Венери» Венера пливе до берега на стулці велетенської мушлі. Черепашка присутня також на фресці з аналогічним сюжетом, яку знайшли в Помпеях. Мушлі морських гребінців були істотною частиною культу богині-матері у Фесті[229].
Цікаві факти
- За останні 4 роки експорт молюсків з України зріс більш як у 10 разів. Так, у 2013 році експортовано 3 тонни равликів, а у 2016 — більше ніж 347[230] тонн. За результатами року равликів експортували в 7 разів більше ніж сала. Середня ціна продукції становить 1 долар за кг. Єдині імпортери — Румунія (25 %) та Литва (75 %). У цих країнах молюсків обробляють і охолодженими відправляють до кінцевих споживачів — Франції, Угорщини, Італії та Польщі.
Див. також
Примітки
- Ponder, Lindberg, 2008, с. 19.
- Rawat, 2010, с. 1.
- Chapman A. D. (2009) Numbers of Living Species in Australia and the World. Australian Biological Resources Study, Canberra. ISBN 978-0-642-56860-1.
- Анистратенко, Анистратенко, 2001, с. 15.
- Rawat, 2010, с. 9.
- Extreme Bivalves. Paleontological Research Institution and its Museum of the Earth. Архівовано з першоджерела 08.01.2014 р.
- O'Shea, S. and Bolstad, K. (2008) Giant Squid and Colossal Squid Fact Sheet. The Octopus News Magazine Online. TONMO.com
- Cowie, R. H., Regnier, C., Fontaine, B., & Bouchet, P. (2017). Measuring the sixth extinction: what do mollusks tell us. Nautilus, 131(1), 3-41.
- Walde A., Hofmann J. B. 1938. Lateinisches etymologisches Wörterbuch. — Heidelberg: Carl Winter's Universitätsbuchhandlung. — S. 103.
- de Vaan M. 2008. Etymological dictionary of Latin and the other Italic languages. — Leiden — Boston: Brill. — P. 386.
- Little L., Fowler H. W., Coulson J., Onions C. T. (editors) 1964. Shorter Oxford English Dictionary. — Oxford University press.
- Рябцева Ю. С. Сравнительная конхология эмбрионов живородок Contectiana listeri (Gastropoda: Viviparidae) из проточных и стоячих водоемов Европы // Вісник Черкаського університету. Серія «Біологічні науки». Випуск № 39 (252), 2012. — С. 99—104. ISSN 2076-5835
- Конхиология Архівовано 31 серпня 2014 у Wayback Machine. // Большая Советская Энциклопедия. — 3-е изд. — Москва: Советская Энциклопедия. — Т. 13. — С. 88. — 608 с. — 629 000 экз.
- Ruppert, Fox, Barnes, 2004, с. 300–343.
- Salvini-Plawen, L. V. & Steiner G. 1996. Synapomorphies and symplesiomorphies in higher classification of Mollusca. Origin and evolutionary radiation of Mollusca. — J. Taylor ed., Oxford Univ. Press. — P. 29—51.
- Giribet, G., Okusu, A., Lindgren, A. R., Huff, S. W., Schrödl, M., and Nishiguchi, M. K. (2006). «Evidence for a clade composed of molluscs with serially repeated structures: Monoplacophorans are related to chitons». Proceedings of the National Academy of Sciences of the United States of America, 103 (20): 7723—7728. DOI:10.1073/pnas.0602578103. PMID 16675549.
- Hayward P. J. 1996. Handbook of the Marine Fauna of North-West Europe. — Oxford University Press. — P. 484—628. — ISBN 0-19-854055-8
- Rawat, 2010, с. 4.
- Rawat, 2010, с. 142.
- Rawat, 2010, с. 5.
- Wilbur, Karl M.; Trueman, E. R.; Clarke, M. R. (eds.) 1985. The Mollusca, 11. Form and Function. — New York: Academic Press. — P. 4. — ISBN 0-12-728702-7
- Ruppert, Fox, Barnes, 2004, с. 284–291.
- Shigeno, S.; Sasaki, T.; Moritaki, T.; Kasugai, T.; Vecchione, M.; Agata, K. (2008). «Evolution of the cephalopod head complex by assembly of multiple molluscan body parts: Evidence from Nautilus embryonic development». Journal of Morphology, 269 (1): 1—17. DOI:10.1002/jmor.10564. PMID 17654542
- Hayward P. J. Handbook of the Marine Fauna of North-West Europe. — Oxford University Press, 1996. — P. 484—628. — ISBN 0-19-854055-8
- Rawat, 2010, с. 6.
- Шарова, 2002, с. 277—279.
- Rawat, 2010, с. 8.
- Шарова, 2002, с. 330.
- Шарова, 2002, с. 291.
- Стаття «Моллюски[недоступне посилання з липня 2019]» з Великої радянської енциклопедії (рос.). Перевірено 6 вересня 2013
- Пойкилосмотические животные Архівовано 31 серпня 2014 у Wayback Machine. — стаття з Великої радянської енциклопедії
- Осморегуляция Архівовано 30 липня 2014 у Wayback Machine. — стаття з Великої радянської енциклопедії
- Dillon, Robert T. The Ecology of Freshwater Molluscs. — Cambridge University Press, 2000. — 509 p.
- Kidney-like organ helps freshwater snail osmoregulate
- Saxena, 2005, с. 2.
- Асеев Н. Эфферентная копия активности нейронов в пищевом поведении виноградной улитки Helix pomatia L.
- Пименова, Пименов, 2005, с. 121—122.
- Пименова, Пименов, 2005, с. 122.
- Пименова, Пименов, 2005, с. 119.
- Шарова, 2002, с. 324.
- Пименова, Пименов, 2005, с. 124.
- Hartfield, P. and E. Hartfield. (1996). Observations on the conglutinates of Ptychobranchus greeni (Conrad, 1834) (Mollusca: Bivalvia: Unionoidea). American Midland Naturalist, 135(2): 370—375.
- Шарова, 2002, с. 312.
- Пименова, Пименов, 2005, с. 130.
- Encyclopedia of Life: Introduction to Mollusks
- Ridgway, I. D.; Richardson, C. A.; Austad, S. N. 2011. «Maximum Shell Size, Growth Rate, and Maturation Age Correlate With Longevity in Bivalve Molluscs». J Gerontol A Biol Sci Med Sci., 66 А (2): 183—190. DOI:10.1093/gerona/glq172
- Стаття «Продолжительность жизни Архівовано 31 серпня 2014 у Wayback Machine.» з Великої радянської енциклопедії (російською мовою). Перевірено 8 вересня 2013
- Rawat, 2010, с. 7.
- Hogan, Michael C. Марианская впадина Архівовано 25 липня 2013 у WebCite. Загадки нашей планеты. zoopage.ru (23 серпня 2010). (рос.) Перевірено 6 вересня 2013
- Шарова, 2002, с. 279.
- Шарова, 2002, с. 283.
- Шарова, 2002, с. 284.
- Шарова, 2002, с. 321.
- Шарова, 2002, с. 286.
- Догель, 1981, с. 470.
- Шарова, 2002, с. 288.
- Шарова, 2002, с. 298.
- Пименова, Пименов, 2005, с. 127.
- Nekhaev, I. O. 2011. Two species of parasitic molluscs new for Russian seas. Ruthenica, 21 (1): 69—72
- Шарова, 2002, с. 304.
- Шарова, 2002, с. 295.
- Догель, 1981, с. 481.
- Шарова, 2002, с. 315.
- Пименова, Пименов, 2005, с. 134.
- Шарова, 2002, с. 316.
- Lützen, J.; Berland, B.; Bristow, G. A. (2011). «Morphology of an endosymbiotic bivalve, Entovalva nhatrangensis (Bristow, Berland, Schander & Vo, 2010) (Galeommatoidea)». Molluscan Research, 31 (2): 114—124.
- Шарова, 2002, с. 322.
- Догель, 1981, с. 498.
- Шарова, 2002, с. 332.
- Шарова, 2002, с. 333.
- Steneck, R. S.; Watling, L. 1982. Feeding capabilities and limitation of herbivorous molluscs: A functional group approach. Marine biology, 68 (3): 299—319. DOI:10.1007/BF00409596
- Tendal, Ole Secher. «Xenophyophores (Protozoa, Sarcodina) in the diet of Neopilina galatheae (Molluscs, Monoplacophora)». Galathea, 16: 95—98.
- Gele, H. 1989. Diet of some Antarctic nudibranchs (Gastropoda, Opisthobranchia, Nudibranchia). Marine biology, 100 (4): 439—441. DOI:10.1007/BF00394819
- Current Organic Chemistry
- Lambert W. J. «Coexistence of Hydroid Eating Nudibranchs: Do Feeding Biology and Habitat Use Matter?». DOI:10.2307/1542096
- Crown of thorns starfish (Acanthaster planci). Архів оригіналу за 5 жовтня 2013. Процитовано 1 жовтня 2013.
- Солемии — стаття з Біологічного енциклопедичного словника.
- Пименова, Пименов, 2005, с. 135.
- Jensen K. R. (2007). «Biogeography of the Sacoglossa (Mollusca, Opisthobranchia) Архівовано 5 жовтня 2013 у Wayback Machine.». Bonner zoologische Beiträge, 55 (2006)(3—4): 255—281.
- Шарова, 2002, с. 276.
- Шарова, 2002, с. 318.
- Barker, G. M. 2004. Natural Enemies of Terrestrial Molluscs. CABI, 644 p. ISBN 0-85199-319-2
- Derby, Charles D. 2007. Escape by Inking and Secreting: Marine Molluscs Avoid Predators Through a Rich Array of Chemicals and Mechanisms. Biol. Bull., 213 (3): 274—289.
- Thorp, J. H.; Delong, M. D.; Casper1, A. F. (1998). «In situ experiments on predatory regulation of a bivalve mollusc (Dreissena polymorpha) in the Mississippi and Ohio Rivers». Freshwater Biology, 39 (4): 649—661. DOI:10.1046/j.1365-2427.1998.00313.x.
- Hulscher, J. B. (1982). «The oystercatcher Haematopus ostralegus as a predator of the bivalve Macoma balthica in the Dutch Wadden Sea[недоступне посилання з квітня 2019]». Ardea 70: 89—152.
- Ingolfsson, Agnar; Bruce T. Estrella (1978). «The development of shell-cracking behavior in herring gulls». The Auk, 95 (3): 577—579.
- Hall, K. R. L.; Schaller, G. B. (1964). «Tool-using behavior of the California sea otter». Journal of Mammalogy 45 (2): 287—298. DOI:10.2307/1376994DOI:
- Fukuyamaa, A. K.; Olivera, J. S. (1985). «Sea star and walrus predation on bivalves in Norton Sound, Bering Sea, Alaska». Ophelia 24 (1): 17-36. DOI:10.1080/00785236.1985.10426616
- Harper, Elizabeth M. (1990). «The role of predation in the evolution of cementation in bivalves[недоступне посилання з квітня 2019]». Palaeontology 34 (2): 455—460.
- Wodinsky, Jerome (1969). «Penetration of the shell and feeding on gastropods by octopus». American Zoologist 9 (3): 997—1010. DOI:10.1093/icb/9.3.997
- Стаття Моллюски з Енциклопедії Кольєра (російською мовою). Перевірено 10 вересня 2013
- Pacific razor clam. California Department of Fish and Game (2001).
- Bourquin, Avril. (2000). Bivalvia: The foot and locomotion Архівовано 24 серпня 2013 у WebCite. The Phylum Mollusca.
- Hodgson, A. N. (1981). «Studies on wound healing, and an estimation of the rate of regeneration, of the siphon of Scrobicularia plana (da Costa)». Journal of Experimental Marine Biology and Ecology 62 (2): 117—128. DOI:10.1016/0022-0981(82)90086-7
- Fleming, P. A.; Muller, D.; Bateman, P. W. (2007). «Leave it all behind: a taxonomic perspective of autotomy in invertebrates». Biological Reviews 82 (3): 481—510. DOI:10.1111/j.1469-185X.2007.00020.x
- Ates, R. M. L. Observations on the symbiosis between Colus gracilis (Da Costa, 1778) (Mollusca: Gastropoda) and Hormathia digitata (O.F. Müller, 1776) (Cnidaria: Actiniaria).
- Farmer, Mark A., Fitt, William K., Trench, Robert K. 2001. Morphology of the Symbiosis Between Corculum cardissa (Mollusca: Bivalvia) and Symbiodinium corculorum (Dinophyceae). Biol. Bull., 200: 336—343.
- Пименова, Пименов, 2005, с. 131.
- Мухин В. А., Третьякова А. С. Биологическое разнообразие: водоросли и грибы. — Ростов-на-Дону: Феникс, 2013. — С. 42—43. — 269 с. — ISBN 978-5-222-20177-0
- Bizarre Bioluminescent Snail: Secrets of Strange Mollusk and Its Use of Light as a Possible Defense Mechanism Revealed
- Haddock, Steven H. D.; Moline, Mark A.; Case, James F. 2010. Bioluminescence in the Sea. Annual Review of Marine Science, 2: 443—493. DOI:10.1146/annurev-marine-120308-081028
- Herring P. J. 1977. Luminescence in cephalopods and fish. Symp. Zool. Soc. Lond., 38: 127—159.
- Ruby E. G., McFall-Ngai M. J. 1992. A squid that glows in the night: development of an animal-bacterial mutualism. J. Bacteriol., 174: 4865—4870.
- Jones B, Nishiguchi M. 2004. Counterillumination in the Hawaiian bobtail squid, Euprymna scolopes Berry (Mollusca: Cephalopoda). Mar. Biol., 144: 1151—1155.
- Nyholm S. V., McFall-Ngai M. 2004. The winnowing: establishing the squid-vibrio symbiosis. Nat. Rev. Microbiol., 2: 632.
- Nyholm S. V., Stewart J. J., Ruby E. G., McFall-Ngai M. J. 2009. Recognition between symbiotic Vibrio fischeri and the haemocytes of Euprymna scolopes. Environ. Microbiol., 11: 483—493.
- Herring P. J., Widder E. A., Haddock S. H. D. 1992. Correlation of bioluminescence emissions with ventral photophores in the mesopelagic squid Abralia veranyi (Cephalopoda: Enoploteuthidae). Mar. Biol., 112: 293—298.
- Robison B. H., Reisenbichler K. R., Hunt J. C., Haddock S. H. D. 2003. Light production by the arm tips of the deep-sea cephalopod Vampyroteuthis infernalis. Biol. Bull., 205: 102—109.
- Bush S. L., Robison B. H., Caldwell R. L. 2009. Behaving in the dark: locomotor, chromatic, postural, and bioluminescent behaviors of the deep-sea squid Octopoteuthis deletron Young, 1972. Biol. Bull., 216: 7—22.
- Robison B. H., Young R. E. 1981. Bioluminescence in pelagic octopods. Pac. Sci., 35: 39—44.
- Herring P. J. 2007. Sex with the lights on? A review of bioluminescent sexual dimorphism in the sea. J. Mar. Biol. Assoc. UK., 87: 829—842.
- Johnsen S, Balser E. J., Fisher E. C., Widder E. A. 1999. Bioluminescence in the deep-sea cirrate octopod Stauroteuthis syrtensis Verrill (Mollusca: Cephalopoda). Biol. Bull., 197: 26—39.
- Viral diseases of molluscs: Abalone viral ganglioneuritis (AVG). Архів оригіналу за 12 лютого 2014. Процитовано 12 вересня 2013.
- Berthe, F. Report about mollusc diseases (назви хвороб дані за назвами паразитів-збудників)
- Jones, J. B. Current trends in the study of molluscan diseases. Diseases in Asian Aquaculture, 7.
- Sigwart J. D, Sutton M. D. (2007) Deep molluscan phylogeny: synthesis of palaeontological and neontological data. Proceedings of the Royal Society B: Biological Scientes, 274 (1624): 2413—2419. DOI:10.1098/rspb.2007.0701
- Догель, 1981, с. 443.
- Haszprunar, G. (2001). «Mollusca (Molluscs)». Encyclopedia of Life Sciences. John Wiley & Sons, Ltd. DOI:10.1038/npg.els.0001598
- Ponder, Winston F. & Lindberg, David R. (eds.) 2008. Phylogeny and Evolution of the Mollusca. Berkeley: University of California Press. p. 481. ISBN 978-0-520-25092-5
- Ruppert, Fox, Barnes, 2004, с. 291–292.
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Низшие целомические животные // Зоология беспозвоночных. Функциональные и эволюционные аспекты = Invertebrate Zoology: A Functional Evolutionary Approach / пер. с англ. Т. А. Ганф, Н. В. Ленцман, Е. В. Сабанеевой; под ред. А. А. Добровольского и А. И. Грановича. — 7-е издание. — М.: Академия, 2008. — Том 2. — 448 с. — ISBN 978-5-7695-2740-1
- Соленогастры // Биологический энциклопедический словарь / Под ред. М. С. Гилярова. — М.: Советская энциклопедия, 1986. — С. 592. — 831 с.
- Властов, Матекин, Зацепин и др., 1988, с. 12.
- Ruppert, Fox, Barnes, 2004, с. 292–298.
- Властов, Матекин, Зацепин и др., 1988, с. 9.
- Ricketts, Edward; Calvin, Jack; Hedgepeth, Joel (1992). Between Pacific Tides (5th edition). Stanford University Press. ISBN 0-8047-2068-1
- Schwabe, E. (2008) A summary of reports of abyssal and hadal Monoplacophora and Polyplacophora (Mollusca). Zootaxa, 1866: 2005—222.
- Vaught, K. C. 1989. A Classification of the Living Mollusca. American Malacologists. ISBN 978-0-915826-22-3
- Ruppert, Fox, Barnes, 2004, с. 367.
- Knop, Daniel. 1996. Giant clams a comprehensive guide to the identification and care of Tridacnid clams. Ettlingen: Dähne Verlag. p. 31. ISBN 3-921684-23-4
- Ruppert, Fox, Barnes, 2004, с. 403–407.
- Introduction to the Scaphopoda
- Ruppert, Fox, Barnes, 2004, с. 300.
- Wells F. E., Walker D. I. & Jones D. S. (eds.) 2003. Food of giants — field observations on the diet of Syrinx aruanus (Linnaeus, 1758) (Turbinellidae) the largest living gastropod. The Marine Flora and Fauna of Dampier, Western Australia. Western Australian Museum, Perth.
- Ruppert, Fox, Barnes, 2004, с. 343.
- Wells, M. J. (1980). «Nervous control of the heartbeat in octopus». Journal of Experimental Biology, 85 (1): 112.
- Wood, J. B. & R. K. O'Dor. 2000. Do larger cephalopods live longer? Effects of temperature and phylogeny on interspecific comparisons of age and size at maturity Архівовано 14 грудня 2004 у Wayback Machine.. Marine Biology 136 (1): 91—99.
- Clarkson, E. N. K. (1998). Invertebrate Palaeontology and Evolution. Blackwell. p. 221. ISBN 0-632-05238-4
- Runnegar, B. and Pojeta, J. (1974). «Molluscan phylogeny: the paleontological viewpoint». Science, 186 (4161): 311—7. DOI:10.1126/science.186.4161.311. Bibcode: 1974Sci…186..311R
- Brusca, R. C. and Brusca, G. J. 2003. Invertebrates. Sinauer Associates. 702 p. ISBN 0-87893-097-3
- Ruppert, Fox, Barnes, 2004, с. 298–300.
- Ruppert, Fox, Barnes, 2004, с. 343–367.
- Ruppert, Fox, Barnes, 2004, с. 367–403.
- Fedonkin, M. A.; Waggoner, B. M. (1997). «The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism». Nature, 388 (6645): 868. DOI:10.1038/42242. Bibcode: 1997Natur.388..868F
- Fedonkin, M. A., Simonetta, A. and Ivantsov, A. Y. (2007). «New data on Kimberella, the Vendian mollusc-like organism (White Sea region, Russia): palaeoecological and evolutionary implications». Geological Society, London, Special Publications, 286: 157—179. DOI:10.1144/SP286.12. Bibcode: 2007GSLSP.286..157F
- Butterfield, N. J. (2006). «Hooking some stem-group „worms“: fossil lophotrochozoans in the Burgess Shale». BioEssays, 28 (12): 1161—1166. DOI:10.1002/bies.20507 PMID 17120226
- Sigwart, J. D. and Sutton, M. D. (2007). «Deep molluscan phylogeny: synthesis of palaeontological and neontological data». Proceedings of the Royal Society: Biology, 274 (1624): 2413—2419. DOI:10.1098/rspb.2007.0701. PMID 17652065. Резюме — у «The Mollusca». University of California Museum of Paleontology.
- Caron, J. B.; Scheltema, A., Schander, C. and Rudkin, D. (2006). «A soft-bodied mollusc with radula from the Middle Cambrian Burgess Shale». Nature, 442 (7099): 159—163. DOI:10.1038/nature04894. PMID 16838013. Bibcode: 2006Natur.442..159C
- Butterfield, N. J. (2008). «An Early Cambrian Radula». Journal of Paleontology, 82 (3): 543—554. DOI:10.1666/07-066.1
- Runnegar, B. and Pojeta, J. (1974). «Molluscan phylogeny: the paleontological viewpoint». Science, 186 (4161): 311—317. DOI:10.1126/science.186.4161.311. PMID 17839855. Bibcode: 1974Sci…186..311R
- Parkhaev, P. Yu. (2007). «The Cambrian 'basement' of gastropod evolution». Geological Society, London, Special Publications, 286: 415—421. DOI:10.1144/SP286.31. Bibcode: 2007GSLSP.286..415P
- Steiner, Michael; Li, Guoxiang; Qian, Yi; Zhu, Maoyan; Erdtmann, Bernd-Dietrich. (2007). «Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China)». Palaeogeography, Palaeoclimatology, Palaeoecology, 254: 67—99. DOI:10.1016/j.palaeo.2007.03.046
- Mus, M. M.; Palacios, T.; Jensen, S. (2008). «Size of the earliest mollusks: Did small helcionellids grow to become large adults?». Geology, 36 (2): 175. DOI:10.1130/G24218A.1
- Landing E., Geyer G., Bartowski K. E. (2002). «Latest Early Cambrian Small Shelly Fossils, Trilobites, and Hatch Hill Dysaerobic Interval on the Quebec Continental Slope». Journal of Paleontology, 76 (2): 287—305. DOI:10.1666/0022-3360(2002)076<0287:LECSSF>2.0.CO;2
- Ruppert, Fox, Barnes, 2004, с. 300—343.
- Frýda J. Paleozoic Gastropoda. In: Phylogeny and evolution of the Mollusca, edited by W. F. Ponder, D. R. Lindberg. California Press, 2008, pp. 239—264. ISBN 0-520-25092-3
- Kouchinsky, A. (2000). «Shell microstructures in Early Cambrian molluscs». Acta Palaeontologica Polonica, 45 (2): 119—150.
- Hagadorn J. W., Waggoner B. M. Proterozoic-Cambrian of the Great Basin and Beyond, Pacific Section SEPM Book 93 Архівовано 1 жовтня 2008 у Wayback Machine., edited by F. A. Corsetti. SEPM (Society for Sedimentary Geology), 2002, pp. 135—150.
- Vickers-Rich, P., Fenton, C. L., Fenton, M. A. and Rich, T. H. 1997. The Fossil Book: A Record of Prehistoric Life. Courier Dover Publications, pp. 269—272. ISBN 0-486-29371-8
- Marshall C. R., and Ward P. D. (1996). «Sudden and Gradual Molluscan Extinctions in the Latest Cretaceous of Western European Tethys». Science, 274 (5291): 1360—1363. DOI:10.1126/science.274.5291.1360
- Pojeta, J. (2000). «Cambrian Pelecypoda (Mollusca)». American Malacological Bulletin, 15: 157—166.
- Schneider, J. A. (2001). «Bivalve systematics during the 20th century». Journal of Paleontology, 75 (6): 1119—1127. DOI:10.1666/0022-3360(2001)075<1119:BSDTC>2.0.CO;2
- Gubanov, A. P., Kouchinsky, A. V. and Peel, J. S. (2007). «The first evolutionary-adaptive lineage within fossil molluscs». Lethaia, 32 (2): 155—157. DOI:10.1111/j.1502-3931.1999.tb00534.x
- Gubanov, A. P., and Peel, J. S. (2003). «The early Cambrian helcionelloid mollusc Anabarella Vostokova». Palaeontology, 46 (5): 1073—1087. DOI:10.1111/1475-4983.00334
- Zong-Jie, F. (2006). «An introduction to Ordovician bivalves of southern China, with a discussion of the early evolution of the Bivalvia». Geological Journal, 41 (3—4): 303—328. DOI:10.1002/gj.1048
- Malinky J. M. (2009). «Permian Hyolithida from Australia: The Last of the Hyoliths?». Journal of Paleontology, 83 (1): 147—152.
- Rawat, 2010, с. 11.
- The Mollusca. University of California Museum of Paleontology.
- Goloboff, Pablo A.; Catalano, Santiago A.; Mirande, J. Marcos; Szumik, Claudia A.; Arias, J. Salvador; Källersjö, Mari; Farris, James S. 2009. Phylogenetic analysis of 73 060 taxa corroborates major eukaryotic groups. Cladistics, 25 (3): 211—230. DOI:10.1111/j.1096-0031.2009.00255.x
- Introduction to the Lophotrochozoa. University of California Museum of Paleontology.
- Henry, J.; Okusu, A.; Martindale, M. 2004. The cell lineage of the polyplacophoran, Chaetopleura apiculata: variation in the spiralian program and implications for molluscan evolution. Developmental biology, 272 (1): 145—160. DOI:10.1016/j.ydbio.2004.04.027
- Jacobs, D. K.; Wray, C. G.; Wedeen, C. J.; Kostriken, R.; Desalle, R.; Staton, J. L.; Gates, R. D.; Lindberg, D. R. 2000. Molluscan engrailed expression, serial organization, and shell evolution. Evolution & Development, 6 (2): 340—347. DOI:10.1046/j.1525-142x.2000.00077.x
- Porter, S. M. 2007. Seawater chemistry and early carbonate biomineralization. Science, 5829 (316): 1301—1302. ISSN 0036-8075. DOI:10.1126/science.1137284
- Вид Achatinella mustelina на сайті Червоного списку Міжнародного союзу охорони природи (англ.). Перевірено 8 вересня 2013
- Офіційний сайт Червоного списку Міжнародного союзу охорони природи (IUCN Red List of Threatened Species. Version 2013.1.) (англійською мовою). Перевірено 8 вересня 2013
- Червона книга України. Тваринний світ / за ред. І. А. Акімова. — Київ: Глобалконсалтинг, 2009. — С. 288—307. — 600 с. — ISBN 978-966-97059-0-7
- Kay, Alison E. 1995. The Conservation Biology of Molluscs. International Union for Conservation of Nature and Natural Resources. ISBN 2-8317-0053-1
- Mannino, M. A. and Thomas, K. D. (2002). «Depletion of a resource? The impact of prehistoric human foraging on intertidal mollusc communities and its significance for human settlement, mobility and dispersal». World Archaeology, 33 (3): 452—474. DOI:10.1080/00438240120107477
- Garrow, J. S., Ralph, A. and James, W. P. T. 2000. Human Nutrition and Dietetics. — Elsevier Health Sciences. — 370 p. — ISBN 0-443-05627-7
- FAO 2010: Fishery and Aquaculture Statistics
- Importing fishery products or bivalve molluscs. United Kingdom: Food Standards Agency. (англійською мовою). Перевірено 8 вересня 2013
- Шарова, 2002, с. 301.
- Шарова, 2002, с. 317.
- Tewfik, A. (1991). An assessment of the biological characteristics, abundance, and potential yield of the queen conch (Strombus gigas L.) fishery on the Pedro Bank off Jamaica. Thesis submitted in partial fulfilment of the requirements for the Degree of Masters of Science (Biology). Acadia University, Canada.
- Шарова, 2002, с. 334.
- Jones, J. B. and Creeper, J. (2006). «Diseases of Pearl Oysters and Other Molluscs: a Western Australian Perspective». Journal of Shellfish Research, 25 (1): 233—238. DOI:10.2983/0730-8000(2006)25[233:DOPOAO]2.0.CO;2
- Иванов А. В. Раковина Архівовано 31 серпня 2014 у Wayback Machine. // Большая Советская Энциклопедия / Под ред. А. М. Прохорова. — 3-е изд. — М.: Советская Энциклопедия, 1975. — Т. 21. — С. 447. — 640 с. — 630 000 экз.
- Gulick, C. B. Athenaeus, The Deipnosophists. — Cambridge, Mass.: Harvard University Press, 1941. — ISBN 0-674-99380-2
- Ziderman, Irving I. 3600 Years of Purple-Shell Dyeing: Characterization of Hyacinthine Purple (Tekhelet). Advances in Chemistry, Vol. 212.
- Webster's Third New International Dictionary (Unabridged). 1976. G. & C. Merriam Co., p. 307.
- Turner, R. D. and Rosewater, J. (1958). «The Family Pinnidae in the Western Atlantic». Johnsonia, 3 (38): 294.
- Hogendorn, J. and Johnson, M. 2003. The Shell Money of the Slave Trade. Cambridge University Press. ISBN 052154110
- Maurer, B. (2006). «The Anthropology of Money Архівовано 19 липня 2011 у Wayback Machine.». Annual Review of Anthropology, 35: 15—36. DOI:10.1146/annurev.anthro.35.081705.123127
- http://zdorovia.com.ua/zhinochezdorovya/ravlikoterapiya-nove-slovo-v-kosmetologii.html «Равликотерапія» — нове слово в косметології
- Краснов И. С. Гигантские улитки-ахатины. Опыт успешного содержания и разведения в домашних условиях. — 2007. — 48 с. — ISBN 5-98435-484-5
- Золотницкий Н. Ф. Аквариум любителя. — М.: ТЕРРА, 1993. — С. 570.
- Вершинина Т. А. Беспозвоночные в аквариуме. Обзор видов. Жизнь в природе. Содержание в аквариуме. — 2008. — ISBN 978-5-9934-0170-6
- Dakin, N. (1993). The Book of the Marine Aquarium. Tetra Press. 400 p. ISBN 1-56465-102-9
- Concar, D. (1996). «Doctor snail—Lethal to fish and sometimes even humans, cone snail venom contains a pharmacopoeia of precision drugs Архівовано 3 травня 2008 у Wayback Machine.». New Scientist.
- Haddad, V.(junior), de Paula Neto, J. B. and Cobo, V. J. (2006). «Venomous mollusks: the risks of human accidents by Conus snails (Gastropoda: Conidae) in Brazil». Revista da Sociedade Brasileira de Medicina Tropical, 39 (5): 498—500. DOI:10.1590/S0037-86822006000500015
- MacRae, Michael. «Engineering Drugs from Marine Mollusks» // ASME.org. (англійською мовою)
- R. Granit. Award Ceremony Speech. The Nobel Prize in Physiology or Medicine 1963 Sir John Eccles, Alan Hodgkin, Andrew Huxley. Nobelprize.org. Nobel Media AB 2014. Web. 29 Oct 2017(англ.)
- Abramson, Charles I.; Shuranova, Zhanna Petrovna (1996). 4. Innate and aquired behaviour of Mollusks. Russian Contributions to Invertebrate Behavior. Greenwood Publishing Group. с. 79–. ISBN 978-0-275-94525-1.(англ.)
- «Physiology or Medicine for 2000 — Press Release». Nobelprize.org. Nobel Media AB 2014. Web. 29 Oct 2017(англ.)
- Rawat, 2010, с. 11-12.
- Московская Н. Раковины мира. История, коллекционирование, искусство. — Аквариум-Принт, Харвест, 2007. — 256 с. — ISBN 978-5-98435-602-2
- Barker, G. M. 2002. Molluscs As Crop Pests. CABI Publications. ISBN 0-85199-320-6
- Civeyrel, L., and Simberloff, D. (1996). «A tale of two snails: is the cure worse than the disease?». Biodiversity and Conservation, 5 (10): 1231—1252. DOI:10.1007/BF00051574
- Benson, Amy. New Zealand Mudsnail Potamopyrgus antipodarum. Southeast Ecological Science Center, U. S. Geological Survey.
- Levri, E. P.; Kelly, A. A.; Love, E. 2007. The invasive New Zealand mud snail (Potamopyrgus antipodarum) in Lake Erie. Journal of Great Lakes Research, 33 (1): 1—6.
- Great Smoky Mountains National Park — Biological threats. Служба національних парків США. (англ.) Перевірено 10 вересня 2013
- Карпов Н. Корабельный червь. «Земля и люди», січень-березень 2009. (російською мовою). Перевірено 10 вересня 2013
- Шарова, 2002, с. 319.
- Alafaci, A. Blue ringed octopus. Australian Venom Research Unit.
- Anderson R. C. 1995. Aquarium husbandry of the giant Pacific octopus. Drum and Croaker, 26: 14—23.
- Brazzelli, V., Baldini, F., Nolli, G., Borghini, F., and Borroni, G. (1999). «Octopus apollyon bite». Contact Dermatitis, 40 (3): 169—170. DOI:10.1111/j.1600-0536.1999.tb06025.x PMID
- Anderson R. C. (1999). «An octopus bite and its treatment». The Festivus, 31: 45—46.
- Williamson, J. A., Fenner, P. J., Burnett, J. W., and Rifkin, J. 1996. Venomous and Poisonous Marine Animals: A Medical and Biological Handbook. — UNSW Press. — P. 65—68. — ISBN 0-86840-279-6
- Livett, B. Cone Shell Mollusc Poisoning, with Report of a Fatal Case Архівовано 1 грудня 2012 у WebCite. Department of Biochemistry and Molecular Biology, University of Melbourne.
- Wekell, John C.; Hurst, John; Lefebvre, Kathi A. (2004). «The origin of the regulatory limits for PSP and ASP toxins in shellfish[недоступне посилання з квітня 2019]». Journal of Shellfish Research, 23 (3): 927—930.
- Brown, D. S. Freshwater Snails of Africa and Their Medical Importance. — CRC Press, 1994. — P. 305. — ISBN 0-7484-0026-5
- Пименова, Пименов, 2005, с. 126.
- Кракен. — Три нотатки з видання «Санктпетербургский вестник» за 1779 р., ч. 3., січень, с. 137—140.
- Off-line інтерв'ю Бориса Стругацького від 02.07.2010 року. Subscribe.Ru. (рос.) Перевірено 10 вересня 2013
- Snails in Popular Culture. Infoqis Publishing, Co. (англ.) Перевірено 10 вересня 2013
- «Улитка». Міфологія. (рос.) Перевірено 10 вересня 2013
- de Vries (Ad.) Dictionary of Symbols and Imagery. — Amsterdam: North-Holland Publishing Company, 1976. — P. 430. — ISBN 0-7204-8021-3
- Трофимов, Д. «Пространственные образы рассказа „Kew gardens“ Вирджинии Вульф Архівовано 26 вересня 2013 у Wayback Machine.». Сайт Наталії Пахсарьян. (рос.) Перевірено 10 вересня 2013
- Мифология — Морская раковина: греческая мифология. (рос.) Перевірено 10 вересня 2013
- Україна експортувала равликів в 7 разів більше, ніж сала — Економічна правда
Джерела та література
Українською
- Гураль-Сверлова Н. В., Гураль Р. І. Визначник наземних молюсків України. — Львів : Державний природознавчий музей НАН України, 2012. — 216 с. — ISBN 978-966-02-6569-1.
- Г. Й. Щербак, Д. Б. Царичкова, Ю. Г. Вервес. Зоологія безхребетних. — Київ : Либідь, 1997. — Т. 3. — 352 с. — ISBN 5-325-00662-2.
- Г. Й. Щербак, Д. Б. Царичкова. Зоологія безхребетних. — Друге видання. — Київ : Київський університет, 2008. — 620 с. — ISBN 978-966-439-075-7.
Російською
- Анистратенко В. В., Анистратенко О. Ю. Фауна Украины: в 40 томах. — Киев : Велес, 2001. — Т. 29. Моллюски. Вып. 1. Кн. 1: Класс Панцирные или Хитоны, класс Брюхоногие — Cyclobranchia, Scutibranchia и Pectinibranchia (часть). — 240 с. — ISBN 966-02-2119-3.
- Анистратенко В. В., Халиман И. А., Анистратенко О. Ю. Моллюски Азовского моря. — Киев : Наукова думка, 2011. — 172 с. — ISBN 978-966-00-1112-0.
- Балашов И. А. Фауна Украины. Том 29. Моллюски. Вып. 5. Стебельчатоглазые (Stylommatophora). — Киев : Наукова думка, 2016. — 592 с. — ISBN 978-966-00-1556-2.
- Балашов И. Охрана наземных моллюсков Украины. — Киев : Институт зоологии НАН Украины, 2016. — 272 с. — ISBN 978-966-02-7841-7.
- Властов Б. В., Матекин П. В., Зацепин В. И., Филатова З. А., Шилейко А. А. и др. Тип Моллюски (Mollusca) // Жизнь животных: в 7 томах / под ред. Р. К. Пастернак, гл. ред. В. Е. Соколов. — 2-е изд. — М. : Просвещение, 1988. — Т. 2. Моллюски. Иглокожие. Погонофоры. Щетинкочелюстные. Полухордовые. Хордовые. Членистоногие. Ракообразные. — С. 5—145. — ISBN 5-09-000445-5.
- Догель В. А. Зоология беспозвоночных. — 7-ое изд. — М. : Высшая школа, 1981. — 614 с.
- Малахов В. В., Медведева Л. А. Эмбриональное развитие двустворчатых моллюсков. — М. : Наука, 1991. — 136 с.
- Пименова И. Н., Пименов А. В. Зоология беспозвоночных. Теория. Задания. Ответы. — Саратов : Лицей, 2005. — 288 с. — ISBN 5-8053-0308-6.
- Скарлато О. А. Двустворчатые моллюски умеренных вод северо-западной части Тихого океана. — Л. : Наука, 1981. — 480 с.
- Шарова И. Х. Зоология беспозвоночных. — М. : Владос, 2002. — 592 с. — ISBN 5-691-00332-1.
Англійською
- Cameron R. Slugs and snails. — London : HarperCollins Publishers, 2016. — 508 с. — ISBN 978-0-00-711301-9.
- Ponder, Winston F. & Lindberg, David R. Phylogeny and Evolution of the Mollusca. — USA : University of California Press, 2008. — 469 p. — ISBN 9780520250925.
- Rawat, R. Anatomy of Mollusca. — New Delhi, India : International scientific publishing academy, 2010. — 260 p. — ISBN 9788182930285.
- Ruppert E. E., Fox R. S., Barnes R. D. Invertebrate Zoology. — 7. — Brooks / Cole, 2004. — ISBN 0-03-025982-7.
- Saxena, A. Text Book Of Mollusca. — New Delhi, India : Discovery Publishing House, 2005. — 528 p. — ISBN 9788183560177.
- The biology of terrestrial molluscs / ed. Barker G. M. — Hamilton : CABI Publishing, 2001. — 558 с. — ISBN 0-85199-318-4.
Посилання
- Просвітницька інтернет-програма «Молюски» лабораторії малакології Державного природознавчого музею.
- Малакологічний фонд Державного природознавчого музею.
- Класифікація типу Mollusca на сайті «World Register of Marine Species»(англ.)
- Інформація про викопних молюсків на сайті «Paleobiology Database»(англ.)
|
|
Ця стаття належить до вибраних статей Української Вікіпедії. |