Гідра (рід)
Гі́дра (Hydra) — рід радіально-симетричних хижих тварин простої будови, що живуть у прісній воді.
? Гідра | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||
| Біологічна класифікація | ||||||||||||||||
| ||||||||||||||||
|
Hydra americana | ||||||||||||||||
Посилання | ||||||||||||||||
| ||||||||||||||||
Загальні відомості
Гідри належать до ряду антомедузи класу гідроїдні; вони трапляються в більшості не надто забруднених прісноводних ставків, озер та річок у помірних та тропічних широтах. Звичайно гідри досягають розміру кількох міліметрів (від 1 до 20, при повністю витягнутому тілі) завдовжки. З огляду на відносно невеличкий розмір найкращим інструментом для їхнього дослідження є мікроскоп. У біології гідра є одним з класичних піддослідних об'єктів, у першу чергу завдяки її видатній здатності до регенерації.
Тіло гідри схоже на трубку, яка за допомогою просто побудованого базального диска (розширення на аборальному полюсі тіла) кріпиться до субстрату. Базальний диск є липким завдяки роботі одноклітинних залоз, що виділяють клейкий секрет. На протилежному, оральному, полюсі гідр розташований ротовий отвір, оточений кільцем щупалець кількістю від п'яти до дванадцяти. Кожне щупальце, або кніда, несе високоспеціалізовані жалкі́ клітини-кнідоцити, які вражають здобич отруйним паралізуючим секретом; однак мікроскопічна кількість отрути при ураженні людини може в найгіршому випадку призвести до незначного свербіння.
Основним поживним ресурсом гідр є дрібні водні безхребетні, такі як дафнії, циклопи, коловертки, а також найпростіші. Кілька видів гідр розвинули симбіотичні відносини з одноклітинними зеленими водоростями, які живуть у їхніх тканинах. У цьому випадку гідра надає водоростям захисту від хижаків, а водорості постачають гідрі поживні речовини, вироблені у процесі фотосинтезу.
Морфологія
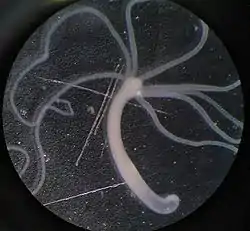
Тіло звичайної гідри має майже правильну циліндричну форму. На одному кінці його міститься рот, оточений віночком із 5—12 тонких довгих щупалець, другий кінець витягнутий у вигляді стебельця з підошвою на кінці. За допомогою підошви гідра прикріплюється до різних підводних предметів, її тіло разом із стебельцем звичайно завдовжки до 7 мм, проте щупальця здатні витягуватися на кілька сантиметрів.
Гідра має променеву симетрію. Якщо вздовж тіла гідри провести уявну вісь, то щупальця розходитимуться від цієї осі в усі боки, як промені від джерела світла. Звисаючи з будь-якої водяної рослини, гідра постійно похитується з боку в бік і повільно водить щупальцями, чатуючи на здобич. Жертва може з'явитися з будь-якого боку, тому променеподібно розставлені щупальця найкраще відповідають такому способові полювання. Променева симетрія загалом характерна, як правило, для тварин, що ведуть прикріплений спосіб життя.
Тіло гідри складається з двох шарів клітин (похідних ектодерми та ентодерми), між якими наявна мезоглія, що являє собою желеподібну неструктуровану речовину; таким чином, як і всі інші гідроїдні поліпи, гідри належать до двошарових тварин (Diploblasta). Зовнішній тканинний шар гідри називається епідермісом, а внутрішній — гастродермісом. Обидва ці шари складаються з відносно просто побудованих клітин.
Нервова система являє собою нервову сітку (дифузний плексус) і є найпростішим з існуючих варіантів нервової системи багатоклітинних тварин. Гідри не мають чітко виділеного мозку та м'язів. Нервові клітини сполучають чутливі фоторецептори та механічні рецептори, що розкидані по зовнішній оболонці та щупальцях гідри.
Дихання здійснюється шляхом дифузії через епідерміс; також цим шляхом здійснюється виведення частини шкідливих продуктів метаболізму.
Будь-які елементи жорсткого скелета у представників роду Гідра відсутні.
У життєвому циклі гідр, на відміну від більшості інших гідроїдних, відсутня стадія медузи: гідра існує у вигляді поліпу впродовж усього життя.
У 19-му столітті деякі науковці зазначали, що гідра є настільки простим організмом, що її можна протерти через тканину, а потім отримана маса знову складеться у життєздатну тварину; попри те що пізніші досліди не підтвердили цього, ця інформація стала поширеною легендою.
Клітинний склад тіла
Епітеліально-м'язові клітини
Епітеліально-м'язові клітини ектодерми і ентодерми утворюють основну масу тіла гідри. У гідри близько 20 000 епітеліально-м'язових клітин.
Клітини ектодерми мають циліндричну форму епітеліальних частин і формують одношаровий покривний епітелій. До мезоглії прилягають скоротні відростки цих клітин, що утворюють поздовжню мускулатуру гідри.
Епітеліально-м'язові клітини ентодерми спрямовані епітеліальними частинами в порожнину кишки і мають по 2-5 джгутиків, які перемішують їжу. Ці клітини можуть утворювати псевдоніжки, за допомогою яких захоплюють частинки їжі. У клітинах формуються травні вакуолі.
Епітеліально-м'язові клітини ектодерми і ентодерми — дві незалежні клітинні лінії. У верхній третині тулуба гідри вони діляться мітотично, а їхні нащадки поступово зміщуються або в бік гіпостома і щупалець, або в бік підошви. У міру переміщення відбувається диференціювання клітин: так, клітини ектодерми на щупальцях дають клітини жалких ділянок тіла, а на підошві — залізисті клітини, що виділяють слиз.
Залозисті клітини ентодерми
Залозисті клітини ентодерми виділяють в порожнину кишки травні ферменти, які розщеплюють їжу. Ці клітини утворюються з інтерстиціальних клітин. У гідри близько 5000 залозистих клітин.
Інтерстиційні клітини
Між епітеліально-м'язовими клітинами знаходяться групи дрібних, округлих клітин, які називаються проміжними, або інтерстиціальними (i-клітини). У гідри їх близько 15 000. Це недиференційовані клітини. Вони можуть перетворюватися в інші типи клітин тіла гідри, крім епітеліально-мускульних. Проміжні клітини мають всі властивості мультипотентних стовбурових клітин. Доведено, що кожна проміжна клітина потенційно здатна дати як статеві, так і соматичні клітини. Стовбурові проміжні клітини не мігрують, проте їх диференціюються клітини-нащадки здатні до швидких міграціям.
Нервові клітини і нервова система
Нервові клітини утворюють в ектодермі примітивну дифузну нервову систему — розсіяне нервове сплетення (дифузний плексус). У ентодермі є окремі нервові клітини. Всього у гідри близько 5000 нейронів. У гідри є згущення дифузного плексусу на підошві, навколо рота і на щупальцях. За новими даними, у гідри є навколоротове нервове кільце, схоже на нервове кільце, що розташоване на краю парасольки у гідромедуз.
У гідри немає чіткого поділу на чутливі, вставні і моторні нейрони. Одна і та ж клітина може сприймати роздратування і передавати сигнал епітеліальної-м'язовим клітинам. Тим не менш, є два основних типи нервових клітин — чутливі і гангліозні. Тіла чутливих клітин розміщені поперек епітеліального пласту, вони мають нерухомий джгутик, оточений комірцем з мікроворсинок, який виходить у напрямку зовнішнього середовища і здатний сприймати подразнення. Гангліозні клітини розташовані в основі епітеліально-м'язових, з відростками що не виходять у зовнішнє середовище. По морфології більшість нейронів гідри — біполярні або мультиполярні.
У нервовій системі гідри присутні як електричні, так і хімічні синапси. З нейромедіаторів у гідри виявлені дофамін, серотонін, норадреналін, гамма-аміномасляна кислота, глутамат, гліцин і багато нейропептидів (вазопресин, речовина Р та інші).
Жалкі клітини
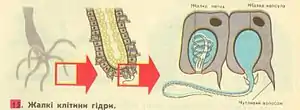
Усе тіло гідри, особливо її щупальця, вкрите великою кількістю жалких клітин.
Жалкі клітини утворюються з проміжних тільки в області тулуба. Спочатку проміжна клітина ділиться 3–5 разів, утворюючи кластер (гніздо) з попередників жалких клітин (кнідобластів), з'єднаних цитоплазматичними містками. Потім починається диференціювання, в ході якої містки зникають. Кнідоцити, що диференціювались, мігрують в щупальця. Жалкі клітини найчисленніші з усіх клітинних типів, їх у гідри близько 55 000.
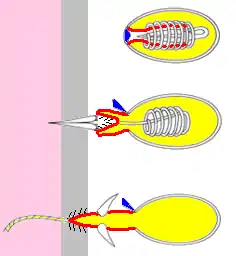
Жалкі клітина має жалку капсулу, заповнену отруйною речовиною. Всередину капсули ввернута жалка нитка. На поверхні клітини знаходиться чутлива нитка, при її подразненні нитка викидається і вражає жертву. Після вистрілювання нитки клітина гине, а з проміжних клітин утворюються нові.
У гідри є чотири типи жалких клітин — стенотели (пенетранти), десмонеми (вольвенти), голотрихи ізорізи (великі глютинанти) і атріхи ізорізи (малі глютинанти). При полюванні першими вистрілюють вольвенти. Їх спіральні жалкі нитки обплутують вирости тіла жертви і забезпечують її утримання. Під дією ривків жертви і викликаної ними вібрації спрацьовують клітини, що мають вищий поріг подразнення — пенетранти. Шипи, наявні у основи їх жалких ниток, заякорюються в тілі жертви, а через порожнисту жалку нитку в її тіло вводиться отрута.
Велика кількість жалких клітин знаходиться на щупальцях, де вони утворюють жалкі батареї. Зазвичай до складу батареї входить одна велика епітеліально-м'язова клітка, в яку занурені жалкі клітини. У центрі батареї знаходиться великий пенетрант, навколо нього — дрібніші вольвенти і глютинанти. Кнідоцити з'єднані десмосомами з м'язовими волокнами епітеліально-м'язової клітини. Великі глютинанти (їх жалка нитка має шипи, але не має, як і у вольвентів, отвору на вершині), мабуть, в основному використовуються для захисту. Малі глютинанти використовуються тільки при пересуванні гідри для міцного прикріплення щупальцями до субстрату. Їх вистрілювання блокується екстрактами з тканин жертв гідри.
Вистрілювання пенетрант гідри було вивчено за допомогою супершвидкісної кінозйомки. Виявилося, що весь процес вистрілювання займає близько 3 мс. У його початковій фазі (до вивертання шипів) швидкість його досягає 2 м/c, а прискорення становить близько 40 000 g (дослідження 1984 року[1][2]); мабуть, це один з найшвидших клітинних процесів з відомих у природі. Першою видимою зміною (менш ніж через 10 мкс після стимуляції) було збільшення об'єму жалкої капсули приблизно на 10%, потім об'єм знижується майже до 50% від початкового. Надалі з'ясувалося, що і швидкість, і прискорення при вистрілювання нематоцист були сильно недооцінені; за даними 2006 року[3], на ранній фазі вистрілювання (викидання шипів) швидкість цього процесу становить 9–18 м/с, а прискорення становить від 1 000 000 до 5 000 000 g. Це дозволяє нематоцисту масою близько 1 нг розвивати на кінчиках шипів (діаметр яких становить близько 15 нм) тиск близько 7 ГПа, що порівняно з тиском кулі на мішень і дозволяє пробивати досить товсту кутикулу жертв.
Статеві клітини та гаметогенез
Як і всім тваринам, гідрам властива оогамія. Більшість гідр роздільностатеві, але зустрічаються гермафродитні лінії гідр. І яйцеклітини, і сперматозоїди утворюються з i-клітин. Вважається, що це особливі субпопуляції i-клітин, які можна відрізнити по клітинних маркерах і які в невеликій кількості присутні у гідр і в період безстатевого розмноження.
При оогенезі ооцити фагоцитують цілі оогонії, а потім кілька ооцитів зливаються, після чого ядро одного з них перетворюється в ядро яйцеклітини, а інші ядра дегенерують. Ці процеси забезпечують швидкий ріст яйцеклітини.
Як нещодавно показано, при сперматогенезі має місце програмована клітинна смерть частини клітин-попередників сперматозоїдів і їх фагоцитування оточуючими клітинами ектодерми [4]
Пересування та рухи
Якщо гідра стривожена або стала об'єктом нападу, її щупальця різко скорочуються, як і все тіло взагалі, зменшуючись в кілька разів і набуваючи приблизно сферичної форми. Завдяки простій будові нервової системи, гідри однаково реагують на будь-який вид подразнення або небезпеки.
Зазвичай гідри ведуть нерухомий спосіб життя, але інколи можуть робити достатньо швидкі рухи, особливо при полюванні. Вони це роблять шляхом згинання та опирання на субстрат ротовим (оральним) полюсом та щупальцями; потім відбувається розгинання з підняттям аборального («підошви») полюса і перенесенням його «через голову» із прикріпленням в новій точці. Ці рухи називаються кульбітами. Такими «перекидами» тварина може пересуватись на відстань до 10 сантиметрів на день. Також гідри можуть пересуватись за рахунок амебоїдних рухів підошви, або просто відкріплюючись від субстрату та пливучи з токами води.
Дихання і виділення
Дихання і виділення продуктів обміну відбувається через всю поверхню тіла тварини. Ймовірно, у виділенні деяку роль відіграють вакуолі, які є в клітинах гідри. Головна функція вакуолей, ймовірно, осморегуляторна; вони виводять надлишки води, які постійно надходять в клітини гідри шляхом осмосу.
Подразливість і рефлекси
Наявність нервової системи дозволяє гідрі здійснювати прості рефлекси. Гідра реагує на механічне подразнення, температуру, наявність у воді хімічних речовин і на ряд інших факторів зовнішнього середовища
Харчування
При харчуванні гідра витягує тіло до максимальної довжини, після чого повільно витягує також і щупальця. Незважаючи на їхню просту будову, щупальця є надзвичайно еластичними і можуть досягати чотирьох-п'яти довжин тіла. Після повного витягування щупальця починають повільно коливатись та згинатись, чатуючи на здобич, що може опинитись в межах їхньої досяжності. Коли потенційна здобич торкається щупальця, нематоцисти «вистрілюють» в неї, а щупальце обвивається навколо. Через приблизно 30 секунд більшість інших щупалець підтягується до здобичі і також обвивають її, «вистрілюючи» при цьому в неї нематоцистами. Через 2-3 хвилини щупальця повністю оточують здобич і починають підтягувати її до відкритого ротового отвора. Ще через приблизно 10 хвилин здобич вміщується в травну порожнину, і починається її перетравлювання. Гідри здатні розтягувати своє тіло таким чином, що всередині травної порожнини вміщується об'єм здобичі, який вдвічі перевищує розмір самої гідри в «голодному» стані. Через 2-3 дні неперетравлені залишки здобичі викидаються назовні через ротовий отвір шляхом швидких скорочень м'язів тіла.
Розмноження
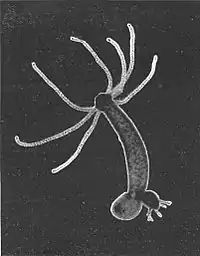
В умовах доброї забезпеченості харчовими ресурсами гідри звичайно розмножуються безстатевим шляхом, продукуючи бруньки на тілі, з котрих виростають аналогічні тварини меншого розміру (див. ілюстрацію), які потім відокремлюються від материнського організму і починають самостійне життя. При наявності кількох таких недосформованих тварин на різних стадіях розвитку їхні травні системи об'єднані і функціонують синхронно, що є аналогічним колоніальним гідроїдам, але це не є справжньою колонією: справжніх довготривалих колоній, де поліпи проводять разом все своє життя, жоден з видів роду Гідра не утворює.
При настанні несприятливих умов, часто пізньої осені, гідри переходять до статевого розмноження. При цьому формуються незапліднені яйця, котрі запліднюються спермою з сперматосаків, які формуються на зовнішній поверхні тіла тварини. Запліднене яйце, залишаючись всередині материнського організму, виділяє щільну зовнішню оболонку, і, після загибелі тварини, ці «сплячі» яйця опиняються в субстраті дна водойми, де перечікують несприятливий період (зиму, посуху тощо). Після настання сприятливих умов з яйця виходить маленька цілком сформована гідра, яка розпочинає новий життєвий цикл.
Гідри є гермафродитами, тобто в одній особині розвиваються як чоловічі (сперматозоїди) так і жіночі статеві клітини (яйцеклітини).
Ріст і регенерація
Міграція та оновлення клітин
У нормі в дорослої гідри клітини всіх трьох клітинних ліній інтенсивно діляться в середній частині тіла і мігрують до підошви, гіпостому і кінчиків щупалець. Там відбувається загибель і злущування клітин. Таким чином, всі клітини тіла гідри постійно оновлюються. При нормальному харчуванні «надлишок» клітин, що діляться, переміщається в бруньки, які зазвичай утворюються в нижній третині тулуба.
Регенераційна здатність
Гідра має дуже високу здатність до регенерації. При розрізуванні упоперек на декілька частин, кожна частина відновлює «голову» і «ногу», зберігаючи вихідну полярність — рот і щупальця розвиваються на тій стороні, яка була ближче до орального кінця тіла, а стеблинка і підошва — на аборальній стороні фрагмента. Цілий організм може відновлюватися з окремих невеликих шматочків тіла (менше 1/100 об'єму тіла тварини), зі шматочків щупалець, а також з суспензії клітин. При цьому сам процес регенерації не супроводжується посиленням клітинних поділів і являє собою типовий приклад морфалаксису.
Гідра може регенерувати із суспензії клітин, отриманих шляхом мацерації. В експериментах показано, що для відновлення головного кінця достатньо агрегату із близько 300 епітеліально-м'язових клітин. Показано, що регенерація нормального організму можлива з клітин одного шару (тільки ектодерми або тільки ентодерми).
Досліди з вивчення регенерації і моделі регенерації
Вже ранні досліди Трамбле показали, що при регенерації зберігається полярність фрагменту. Якщо розрізати тіло гідри поперек на кілька циліндричних фрагментів, то на кожному з них ближче до колишнього орального кінця регенерують гіпостом і щупальця (в експериментальній ембріології гідри закріпився термін «голова» для позначення орального кінця тіла), а ближче до колишнього аборальном полюса — підошва («нога»). При цьому у тих фрагментів, які розташовувалися ближче до «голови», швидше регенерує «голова», а у фрагемнтів розташовувалися ближче до «ноги» — «нога».
Пізніше досліди з вивчення регенерації були вдосконалені в результаті застосування методики зрощування фрагментів різних особин — вирощування химер. Якщо вирізати з бічної сторони тулуба гідри фрагмент і зростити його з тілом іншого гідри, то можливі три результати досвіду:
- фрагмент повністю зливається з тілом реципієнта;
- фрагмент утворює виступ, на кінці якого розвивається «голова» (тобто перетворюється на бруньку);
- фрагмент утворює виступ, на кінці якого утворюється «нога».
З'ясувалося, що відсоток утворень «голів» тим вищий, чим ближче до «голови» донора узятий фрагмент для пересадки і чим далі від «голови» реципієнта він поміщений. Ці та аналогічні досліди призвели до постулювання існування чотирьох речовин-морфогенів, що регулюють регенерацію — активатора та інгібітора «голови» і активатора та інгібітора «ноги». Ці речовини, відповідно до даної моделі регенерації, утворюють концентраційні градієнти: у районі «голови» у нормальної поліпа максимальна концентрація як активатора, так і інгібітора голови, а в районі «ноги» — максимальна концентрація і активатора, і інгібітора ноги.
Ці речовини дійсно були виявлені. Активатор голови — пептид з 11-ти амінокислот (pGlu-Pro-Pro-Gly-Gly-Ser-Lys-Val-Ile-Leu-Phe), активний в пікомолярній концентрації (спочатку 20 мкг активатора були отримані з 200 кг морських актиній) . У людини він присутній в гіпоталамусі і кишечнику і в тій же концентрації володіє нейротрофічною дією. У гідри і ссавців цей пептид має також мітогенну дію і впливає на диференціацію клітин.
Активатор ноги — теж пептид з молекулярною масою, близькою до 1000 Да. Інгібітори голови і ноги — низькомолекулярні гідрофільні речовини небілкової природи. У нормі всі чотири речовини виділяються нервовими клітинами гідри. Активатор голови має більший час напівжиття (близько 4 годин), ніж інгібітор (30 хв) і повільніше дифундує, так як пов'язаний з білком-носієм. Інгібітор голови в дуже низькій концентрації пригнічує виділення активатора, а в 20 разів більшій концентрації — своє власне виділення. Інгібітор ноги також інгібує виділення активатора ноги.
Отримання «безнервових» гідр
При регенерації, як і при рості та безстатевому розмноженні, епітеліально-м'язові клітини діляться самостійно, причому клітини ектодерми і ентодерми — дві незалежні клітинні лінії. Решта типів клітин (нервові, жалкі і залозисті) розвиваються з проміжних. Вбивши проміжні клітини, що діляться, високою дозою радіації або колхіцином, можна отримати «безнервових», або епітеліальних гідр — вони продовжують рости і розмножуватися брунькуванням, але бруньки, що відокремлюються, позбавлені нервових і жалких клітин. Культуру таких гідр вдається підтримувати в лабораторії за допомогою «насильницького» годування.
Відомі також мутантні лінії «безнервових» гідр, у яких немає проміжних клітин і в яких проміжні клітини можуть давати тільки сперматозоїди, але не соматичні клітини, а також мутантні лінії, у яких проміжні клітини гинуть при підвищеній температурі.
«Безнервові» гідри зберігають здатність до регенерації.
Старіння
Давно припускалося, що гідра належить до тих кількох видів тварин, які не старіють і є фактично біологічно безсмертними. Данієл Мартінез (англ. Daniel Martinez) навів докази на користь цієї гіпотези в 1998 році в журналі «Експериментальна геронтологія» (англ. Experimental Gerontology).[5] Ця публікація широко цитувалася як доказ того, що гідра не старіє і як доказ існування нестаріючих організмів загалом. Однак у 2010 році Естеп П. В. (англ. Estep P. W.) опублікував (також в журналі «Експериментальна геронтологія») лист до редактора щодо даних Мартінеза, аргументуючи на користь гіпотези старіння гідри.[6]
Геном
У 2010 році була опублікована частина геному виду Hydra magnipapillata.[7]
Види
- Hydra americana
- Hydra beijingensis
- Hydra canadensis
- Hydra cauliculata
- Hydra circumcincta
- Hydra daqingensis
- Hydra hadleyi
- Hydra harbinensis
- Hydra hymanae
- Hydra iheringi
- Hydra intaba
- Hydra intermedia
- Hydra japonica
- Hydra liriosoma
- Hydra madagascariensis
- Hydra mariana
- Hydra minima
- Hydra mohensis
- Hydra oligactis
- Hydra oregona
- Hydra oxycnida
- Hydra paludicola
- Hydra parva
- Hydra plagiodesmica
- Hydra robusta
- Hydra rutgersensis
- Hydra salmacidis
- Hydra umfula
- Hydra utahensis
- Hydra viridissima
- Hydra vulgaris
Види України
.JPG.webp)
У водоймах України найбільш часто зустрічаються такі види гідр (в даний час багато зоологів виділяють крім роду Hydra ще 2 роди — Pelmatohydra і Chlorohydra):
- Гідра довгостебельчаста (Hydra (Pelmatohydra) oligactis) — велика, з пучком дуже довгих ниткоподібних щупалець, що в 2-5 разів перевищують довжину її тіла;
- Гідра звичайна (Hydra vulgaris) — щупальця приблизно вдвічі довші за тіло, а саме тіло, як і в попереднього виду звужується ближче до підошви;
- Гідра тонка (Hydra attennata) — тіло цієї гідри має вигляд тонкої трубочки рівномірної товщини, а щупальця тільки трішки довші за тіло;
- Гідра зелена (Hydra (Chlorohydra) viridissima) з короткими, але численними щупальцями, трав'янистого-зеленого кольору.
Симбіонти
У так званих «зелених» гідр Hydra (Chlorohydra) viridissima в клітинах ентодерми живуть ендосимбіотичні водорості роду Chlorella — зоохлорелли. На світлі такі гідри можуть тривалий час (понад чотири місяці) обходитися без їжі, в той час як штучно позбавлені симбіонтів гідри без годування гинуть через два місяці. Зоохлорелли проникають в яйцеклітини і передаються потомству трансоваріально. Інші види гідр в лабораторних умовах, інколи вдається заразити зоохлорелами, проте стійкого симбіозу при цьому не виникає.
Саме зі спостережень за зеленими гідрами почав свої дослідження відомий дослідник гідр Авраам Трамбле.
Хижаки і паразити
На гідр можуть нападати мальки риб, для яких опіки жалких клітин, мабуть, досить чутливі: схопивши гідру, мальок зазвичай випльовує її і відмовляється від подальших спроб з'їсти.
На поверхні тіла гідр як паразити або коменсали часто мешкають Kerona polyporum, триходини та інші інфузорії.
До харчування тканинами гідр пристосований гіллястовусі раки з родини хідорид Anchistropus emarginatus.
Тканинами гідр можуть також харчуватися турбелярії мікростомули, які здатні використовувати неперетравлені молоді жалкі клітини гідр як власні захисні клітини — клептокніди.
Історія відкриття та вивчення
Мабуть, вперше описав гідру Антоні ван Левенгук. Детально вивчив харчування, рух і безстатеве розмноження, а також регенерацію гідри Авраам Трамбле, який описав результати своїх дослідів і спостережень в книзі «Мемуари до історії одного роду прісноводних поліпів з руками у формі рогів» (перше видання вийшло французькою мовою в 1744 році). Відкриття Трамбле набуло гучної слави, його досліди обговорювалися у світських салонах і при французькому королівському дворі. Ці досліди спростували пануюче тоді переконання, що відсутність безстатевого розмноження і розвиненої регенерації у тварин — одне з найважливіших їх відмінностей від рослин. Вважається, що вивчення регенерації гідри (досліди А. Трамбле) поклали початок експериментальній зоології. Наукову назву роду згідно з правилами зоологічної номенклатури присвоїв Карл Лінней.
Гідра як модельний об'єкт
В останні десятиліття гідра використовується як модельний об'єкт для вивчення регенерації і процесів морфогенезу. Геном гідри (північноамериканський вид Hydra magnipapillata) частково розшифрований. Є колекції мутантних ліній гідри в Японії і Німеччині. Розроблено методику отримання трансгенних гідр.
Див. також
- Гідроїдні — вибрана стаття
Посилання
- Holstein T., Tardent P. (1984). An ultrahigh-speed analysis of exocytosis: nematocyst discharge. Science 223 (4638): 830–833. PMID 6695186. doi:10.1126/science.6695186.
- Kass-Simon G., Scappaticci A. A. Jr (2002). The behavioral and developmental physiology of nematocysts. Canadian Journal of Zoology 80: 1772–1794. doi:10.1139/Z02-135. Процитовано 25 жовтня 2012.
- Nüchter Timm, Benoit Martin, Engel Ulrike, Özbek Suat, Holstein Thomas W. (2006). Nanosecond-scale kinetics of nematocyst discharge. Current Biology 16 (9): R316–R318. doi:10.1016/j.cub.2006.03.089. Процитовано 25 жовтня 2012.
- Sergey Kuznetsov, Maria Lyanguzowa, Thomas CG Bosch / Role of epithelial cells and programmed cell death in Hydra spermatogenesis / Zoology, Vol. 104, Issue 1, 2001, Pages 25—31 PubMed
- Martinez, D.E. (May 1998). Mortality patterns suggest lack of senescence in hydra. Experimental Gerontology 33 (3): 217–225. PMID 9615920. doi:10.1016/S0531-5565(97)00113-7.
- Estep, P.W. (September 2010). Declining asexual reproduction is suggestive of senescence in hydra: comment on Martinez, D., "Mortality patterns suggest lack of senescence in hydra.". Experimental Gerontology 45 (9): 645–6. PMID 20398746. doi:10.1016/j.exger.2010.03.017.
- Chapman1, Jarrod A.; Kirkness, EF; Simakov, O; Hampson, SE; Mitros, T; Weinmaier, T; Rattei, T; Balasubramanian, PG та ін. (March 2010). The dynamic genome of Hydra. Nature. advance online publication 14 March 2010 (7288): 592–6. PMID 20228792. doi:10.1038/nature08830.
Література
- Gilberson, Lance, Zoology Lab Manual, 4th edition. Primis Custom Publishing. 1999
- Solomon, E., Berg, l., Martin, D., Biology 6th edition. Brooks/Cole Publishing. 2002
- Martinez, D.E. (1998) «Mortality patterns suggest lack of senescence in hydra.» Experimental Gerontology 1998 May;33(3):217-225. Повний текст.
- Степаньянц С. Д., Кузнецова В. Г., Анохин Б. А. Гидра: от Абраама Трамбле до наших дней. — Москва-Санкт-Петербург: Товарищество научных изданий КМК, 2003. — (Разнообразие животных).
- Трамбле А. Мемуары к истории одного рода преноводных полипов с руками в форме рогов. — М.-Л.: Государственное издательство биологической и медицинской литературы, 1937.
- Иофф Н. А. Курс эмбриологии беспозвоночных. — М.: Высшая школа, 1962.
- G. Kass-Simon, A.A. Scappaticci, Jr. The behavioral and developmental physiology of nematocysts. — 2002. — С. 1772—1794. — (Can. J. Zool. Vol. 80).
- Щербак Г. Й., Царичкова Д. Б., Вервес Ю. Г. Зоологія безхребетних — К. : Либідь, 1995.