Ефект Бора
Ефект Бора — це фізіологічне явище, яке вперше було описане 1904 року датським фізіологом Крістіаном Бором. Підвищення кислотності та концентрації вуглекислого газу призводить до зменшення спорідненості гемоглобіну до кисню (див. Криву дисоціації кисню та гемоглобіну).[1] Тобто зі зміною концентрації вуглекислого газу або рН середовища крива дисоціації кисню буде зсуватися. Оскільки вуглекислий газ реагує з водою з утворенням вуглекислоти, збільшення концентрації СО 2 призводить до зниження рН крові [2] і, відповідно, зниження спорідненості гемоглобіну до кисню. І навпаки, зменшення концентрації вуглекислого газу призводить до підвищення рН і збільшує спорідненість гемоглобіну до кисню.
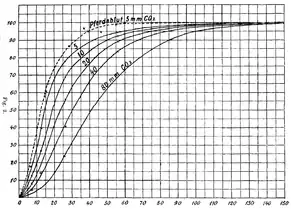

На початку 1900-х років Крістіан Бор був добре відомим своєю роботою в галузі фізіології дихання професором Копенгагенського університету в Данії.[3] Протягом двох десятиріч він вивчав розчинність кисню, вуглекислого газу та інших газів у різних рідинах [4] і провів великі дослідження гемоглобіну та його спорідненості до кисню.[3]1903 року він почав тісно співпрацювати з Карлом Хассельбальхом та Августом Крогом, двома його співробітниками, намагаючись експериментально відтворити роботи Густава фон Хюфнера, використовуючи цільну кров замість розчину гемоглобіну.[1] Хюфнер припустив, що крива зв’язування кисню та гемоглобіну має гіперболічну форму, [5] але після експериментів було визначено, що крива насправді сигмоїдна. Крім того, в процесі побудови численних кривих дисоціації стало очевидним, що високий парціальний тиск вуглекислого газу призводив до зміщення кривих вправо.[4] Подальші експерименти підтвердили це спостереження.
Існує суперечка щодо того, чи насправді Бор виявив зв'язок між СО 2 та спорідненістю до кисню першим, чи російський фізіолог Вериго Броніслав Фортунатович, який нібито виявив цей ефект 1898 року, за шість років до Бора.[6] Хоча це ніколи не було доведено, Вериго фактично опублікував статтю про взаємозв'язок гемоглобіну і СО 2 1892 року.[7] Запропонована ним модель була недосконалою, і Бор жорстоко критикував її у своїх публікаціях.[1]
Також на відкриття цього ефекту претендує Крог, соратник Бора, який винайшов прилад для вимірювання концентрації газу в експериментах [8] і протягом усього життя стверджував, що він сам насправді був першим, хто виявив ефект.[4]
Фізіологічна роль
Ефект Бора підвищує ефективність транспортування кисню через кров. Гемоглобін зв’язується з киснем у легенях, де парціальний тиск кисню високий, а вуглекислого газу низький. А в тканинах, які потребують кисню, концентрація кисню буде низькою, а вуглекислого газу — високою, тому спорідненість гемоглобіну до кисню падає, і він віддає його тканинам.
Коли швидкість метаболізму в тканині зростає, збільшується і вироблення вуглекислого газу. Викидаючись в кров, вуглекислий газ утворює бікарбонат і протони внаслідок такої реакції:

Хоча ця реакція зазвичай протікає дуже повільно, фермент карбоангідраза (який присутній в еритроцитах) різко прискорює утворення бікарбонату.[2] Це призводить до зниження рН крові, що сприяє дисоціації кисню і дозволяє навколишнім тканинам отримувати достатню кількість кисню для задоволення їх потреб. У тканинах, де концентрація кисню є високою, зокрема в легенях, зв’язування кисню змушує гемоглобін вивільняти протони, що вступають у зворотню реакцію з утворенням вуглекислого газу, що видихається. Ці протилежні реакції протонізації та депротонування відбуваються в рівновазі, що призводить до незначної загальної зміни рН крові.
Ефект Бора дозволяє організму адаптуватися до мінливих умов і дає можливість постачати додатковий кисень тканинам, які найбільше потребують цього. Наприклад, під час напруженої роботи м’язів, вони потребують велику кількість кисню, а внаслідок їх роботи утворюється багато CO2 (а, отже, і HCO 3- і H+). CO2 знижує рН крові, що посилює віддачу кисню гемоглобіном. Вуглекислий газ — не єдина молекула, що спричиняє ефект Бора. Якщо м’язові клітини не отримують достатньо кисню для клітинного дихання, вони вдаються до молочнокислого бродіння, внаслідок якого утворюється молочна кислота як побічний продукт. Це підвищує кислотність крові сильніше, ніж лише CO2 (в анаеробних умовах м’язи виробляють молочну кислоту настільки швидко, що рН крові, яка проходить через м’язи, падає до приблизно 7,2), що змушує гемоглобін віддавати на 10% більше кисню.[2]
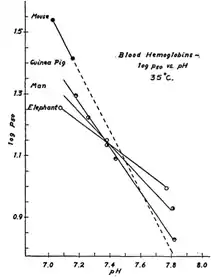

Сила ефекту та розмір тіла
Сила ефекту Бора залежить від розміру організму: величина зростає із зменшенням розміру та ваги. Наприклад, миші мають дуже сильний ефект Бора, що вимагає відносно незначних змін концентрацій H + або CO 2, тоді як слони потребують набагато більших змін концентрації для досягнення набагато слабшого ефект.[9]
Механізм
Алостеричні взаємодії
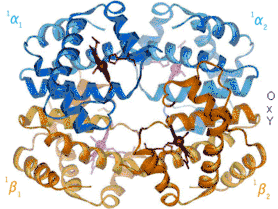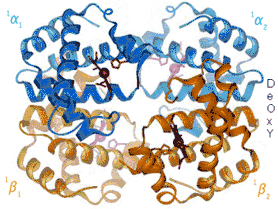
Ефект Бора залежить від алостеричних взаємодій між гемами тетрамеру гемоглобіну, механізм яких був вперше запропонований Максом Перуцем 1970 року.[10] Гемоглобін існує у двох конформаціях: високоафінний R-стан та низькоафінний T-стан. При висока концентрація кисню сприяє стану R і більшому насиченню гемоглобіну.[2] У молекулі гемоглобіну чотири геми працюють узгоджено, кооперативно. Важче за все приєднується перша молекула оксигену, а кожна наступна — легше. Перша молекула оксигену з’єднується з гемом α1-субодиниці, при цьому розриваються два іонні зв’язки між α1— та α2-субодиницями. Субодиниці набувають більшої рухливості, що полегшує приєднання другої молекули оксигену до гему α2-субодиниці. Два α1-α2 іонні зв’язки при цьому також розриваються, що дає можливість останнім гемам зайняти вигідне положення для приєднання оксигену. Третя молекула оксигену приєднується до β1-субодиниці. Один із іонних зв’язків β1-β2 розривається, полегшуючи доступ оксигену до останнього атома заліза гему β2-субодиниці. При цьому розривається останній іонний зв’язок β1-β2. Завантажений оксигеном гемоглобін віддає його спочатку важко, а потім все легше і легше. Така кооперативна поведінка гемоглобіну має фізіологічне значення. Якби геми діяли автономно, то розвантаження оксигену вимагало б 90-кратного перепаду тиску, що неможливо, тому основна маса оксигену не може відриватися від гемоглобіну. Людина задихнулася б навіть в атмосфері кисню.[11] Міоглобін, мономерний білок, не проявляє ефекту Бора.[2]
Стабілізація T-стану
Коли гемоглобін знаходиться у Т-стані, N-кінцеві аміногрупи α-субодиниць та C-кінцевий гістидин β-субодиниць протонуються, що дозволяє цим залишкам брати участь в іонних взаємодіях з карбоксильними групами сусідніх залишків. Ці взаємодії допомагають утримувати гемоглобін у Т-стані. Зниження рН (підвищення кислотності) ще більше стабілізує цей стан, посилюючи протонування. Стабільність стану R зростає зі збільшенням рН. [2]
Діоксид вуглецю може також безпосередньо реагувати з N-кінцевими аміногрупами, утворюючи карбамати, в наступній реакції:

Карбамати найчастіше утворюються в Т-стані, що допомагає стабілізувати цю конформацію. В цьому процесі також виділяються протони, додатково стабілізуючи Т-стан.[2]
Особливі випадки

Морські ссавці
Морські тварини є винятком з правила про зв'язок між розмірами тіла тварини та чутливістю її гемоглобіну до змін рН, який було виявлено 1961 року.[12] Виходячи з їх розміру та ваги, багато морських ссавців мали б мати майже незначний ефект Бора. Однак, не так. У горбатих китів вагою 41000 кілограмів спостерігали значення, що приблизно еквівалентне величині ефекту Бора в 0,57 кг мурчака.[9] Цей надзвичайно сильний ефект Бора є одним із багатьох пристосувань морських ссавців до глибоких, тривалих занурень, оскільки він дозволяє практично всьому зв’язаному з гемоглобіном кисню дисоціювати та забезпечувати довге перебування кита під водою.
Монооксид вуглецю
Інший особливий випадок ефекту Бора має місце, коли присутній монооксид вуглецю. Ця молекула служить конкурентним інгібітором і зв’язується з гемоглобіном, утворюючи карбоксигемоглобін.[13] Спорідненість гемоглобіну до СО приблизно в 210 разів сильніша за спорідненість до O 2, [14] тому він блокує зв'язування O 2 з цією субодиницею. У той же час CO за структурою досить подібний до O 2, тому він сприяє стабілізації R-стану, підвищуючи спорідненість кисню до інших незайнятих субодиниць. Ця комбінація значно зменшує доставку кисню до тканин організму, що робить оксид вуглецю настільки токсичним.[13]
Примітки
- Bohr; Hasselbalch, Krogh. Concerning a Biologically Important Relationship - The Influence of the Carbon Dioxide Content of Blood on its Oxygen Binding.
- Voet, Donald; Judith G. Voet; Charlotte W. Pratt (2013). Fundamentals of Biochemistry: Life at the Molecular Level (вид. 4th). John Wiley & Sons, Inc. с. 189.
- Irzhak, L. I. (2005). Christian Bohr (On the Occasion of the 150th Anniversary of His Birth). Human Physiology (англ.) 31 (3): 366–368. ISSN 0362-1197. doi:10.1007/s10747-005-0060-x.
- Edsall, J. T. (1972). Blood and hemoglobin: the evolution of knowledge of functional adaptation in a biochemical system, part I: The adaptation of chemical structure to function in hemoglobin. Journal of the History of Biology 5 (2). с. 205–257. ISSN 0022-5010. PMID 11610121. doi:10.1007/BF00346659. Процитовано 14 грудня 2020.
- G. Hüfner, "Ueber das Gesetz der Dissociation des Oxyharmoglobins und iiber einige daran sich knupfenden wichtigen Fragen aus der Biologie," [On the Law of the Dissociation of Oxyharmoglobin, and on some important questions arising from biology]. Arch. Anat. Physiol. (in German) (Physiol. Abtheilung) (1890), 1-27.
- Вериго эффект - это... Что такое Вериго эффект? [Verigo effect is... What is the Verigo effect?]. Словари и энциклопедии на Академике (рос.). Процитовано 8 листопада 2016.
- B. Werigo, "Zur Frage uber die Wirkung des Sauerstoffs auf die Kohlensaureausscheidung in den Lungen," [The question about the effect of oxygen on the secretion of carbonic acid in the lungs]. Pflügers Arch. ges. Physiol. (in German), 51 (1892), 321-361.
- A. Krogh, "Apparat und Methoden zur Bestimmung der Aufnahme von Gasen im Blute bei verschiedenen Spannungen der Gase," [Apparatus and methods for the determination of the absorption of gases in the blood at different tensions of the gases]. Skand. Arch. Physiol. (in German), 16 (1904), 390-401.
- Riggs, Austen (1 березня 1960). The Nature and Significance of the Bohr Effect in Mammalian Hemoglobins. The Journal of General Physiology (англ.) 43 (4): 737–752. ISSN 0022-1295. PMC 2195025. PMID 19873527. doi:10.1085/jgp.43.4.737.
- Perutz, Max (15 січня 1998). Science is Not a Quiet Life. World Scientific. ISBN 9789814498517.
- ОКСИГЕМОГЛОБІН. Фармацевтична енциклопедія (укр.). Процитовано 14 грудня 2020.
- Riggs, Austen (1 квітня 1961). Bohr Effect in the Hæmoglobins of Marine Mammals. Nature (англ.) 190 (4770): 94–95. Bibcode:1961Natur.190...94R. PMID 13741621. doi:10.1038/190094a0.
- Hlastala, M. P.; McKenna, H. P.; Franada, R. L.; Detter, J. C. (1976-12). Influence of carbon monoxide on hemoglobin-oxygen binding. Journal of Applied Physiology 41 (6). с. 893–899. ISSN 0021-8987. PMID 12132. doi:10.1152/jappl.1976.41.6.893. Процитовано 14 грудня 2020.
- Hall, John E. (2010). Guyton and Hall Textbook of Medical Physiology (12th ed.). Philadelphia, Pa: Saunders/Elsevier. с. 502. ISBN 978-1416045748.