Жаба їстівна
Їстівна або зелена водна жаба[1] (Pelophylax esculentus) — вид жаб роду Зелена жаба, дуже звичайний в Європі, розмірами від 5 до 9 см (самки) або від 6 до 11 см (самці). Є природним гібридом видів ставкової і озерної жаб, що сформувався близько 5000 років тому. Представники виду широко використовуються в їжу, особливо у Франції.
? Їстівна жаба | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||
| Біологічна класифікація | ||||||||||||||||
| ||||||||||||||||
| Pelophylax esculentus (Linnaeus, 1758) | ||||||||||||||||
 | ||||||||||||||||
Посилання | ||||||||||||||||
| ||||||||||||||||

Історія таксономічного статусу
З'ясування таксономічного статусу їстівних жаб пов'язане з працями польського зоолога Л. Бергера, яким були вивчені дані з гібридизації та мінливості зовнішньоморфологічних ознак. Досліджувалися жаби, що їх традиційно відносили на той час до різних підвидів одного й того ж самого виду: R. esculentus esculentus та R. esculentus lessonae. Результатом праці став висновок, що ці таксони мають розглядатися як самостійні біологічні види. Згодом цей самий автор вказав на гібридогенне походження їстівної жаби.
Подальший розвиток таксономічних досліджень амфібій сформував думку про те, що озерна, ставкова та їстівна жаби входять до роду Pelophylax. Попри те, що гібридогенний таксон звичайно підтримує своє існування шляхом схрещування з одним з батьківських видів, популяції, що складаються лише з представників P. esculentus (без озерної та ставкової жаб) відомі у Данії, Швеції, Німеччині, Польщі, Нідерландах, Франції, Чехії, Словаччині, Росії. Виникнення гібридогенного таксона пов'язують, найбільш вірогідно, з останнім (вюрмським) зледенінням, що дає можливість оцінити вік цих амфібій близько 10 тис. років.
Походження третього таксона комплексу зелених, або водяних жаб, як вже зазначалося, пов'язане з гібридизаційними процесами між двома батьківськими видами: озерною та ставковою жабами. Очевидно, це зумовлює частковий збіг контурів ареалу цього виду з ареалом ставкової жаби у європейській частині її поширення, хоча на заході представники цієї групи досягають Іспанії, на півночі захоплюють майже всю північну частину узбережжя Європи (інтродукована в Англії), на сході доходять до середньої та верхньої течії Волги, на півдні — до Австрії, північної Болгарії, в Україні — до пониззя Дніпра та дельти Дунаю.
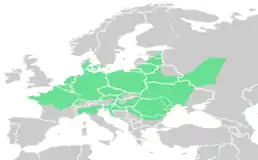
Ареал розповсюдження
В Україні їстівна жаба Р. esculentus живе на більшій частині території, проникаючи на південь у степову зону долинами великих річок.
У виборі місць пробування їстівна жаба тяжіє до біотопів, типовіших для ставкової жаби — ставків, стариць річок, мілководним каналам, невеликим озерам тощо. Водойм у суцільних лісонасадженнях або великих річок зазвичай уникає.
Чисельність
Матеріали з чисельності Р. esculentus перед з'ясуванням специфіки таксономічних взаємовідносин у групі характеризували найчастіше озерну чи ставкову жабу, або ж увесь комплекс у цілому. Однак навіть після детального розв'язання питань, пов'язаних зі специфікою цієї групи, сучасна ідентифікація їстівної жаби при польових дослідженнях, без їх відлову та лабораторного вивчення, додає певних труднощів, і достовірність результатів візуальних обліків чисельності часто залежить від кваліфікації дослідника та його досвіду роботи. У зв'язку з цим далі наводяться відомості про чисельність в основному за частотою трапляння Р. esculentus у загальній вибірці зелених жаб після її вилучення з природи.
Так, в одній популяції цього комплексу у Київській області з 95 особин їстівні жаби становлять 8,4 і 10,5 %, з 77 тварин іншої — 14,3 та 18,2 %, з 46 амфібій третьої — 45,7 та 15,2 % (відповідно F1 та F2).
У трьох вибірках цього комплексу (132 особини) з околиць Ужгорода (Закарпатська обл.) у перших двох були особини виключно гібридогенного таксона, а у третій з 7 тварин 1 також відносилась до Р. esculentus, а 6 — до Р. ridibundus.
Серед 46 зелених жаб з околиць Луцька (Волинська обл.), близько 24 % були Р. esculentus (решта була представлена озерною жабою).
В українській частині дельти Дунаю з 57 особин цього комплексу навесні 1992 р. близько 45 % складали їстівні жаби, восени 1994 р. з 33 жаб — 15 %. В Угорщині у R-E популяційній системі (R + 3nE — самці) з 111 особин 52 жаби належали до Р. esculentus (50 триплоїдних самців, одна диплоїдна самиця та один диплоїдний самець; решта 32 самці та 27 самиць — P. ridibundus).
У різних регіонах Росії у популяціях з Р. esculentus останні трапляються у них з частотою 5 % (Івановська обл.), 7 % (Нижньогородська обл.), 8 % (Удмуртська республіка) або ж (Центрально-Чорноземний регіон) 21 % (Borkin et al., 2002). У Казані (Татарстан, Росія) з 1287 особин 389 були представлені їстівними жабами, решта 618 та 280 — відповідно озерною та ставковою жабами.
Зовнішній вигляд


Жаби гібридогенного таксона, за своїм зовнішнім виглядом схожі з батьківськими видами: Р. ridibundus та Р. lessonae. Це амфібії середніх розмірів, довжина їхнього тулуба може досягати до 100 мм. В забарвленні спинного боку тіла звичайні зелені тони, хоча в частини особин у популяції можуть переважати брунатні відтінки. Візерунок зазвичай представлений різними сполученнями спинної смуги та плям, або ж їх відсутністю. На спині вздовж хребта зазвичай присутня смужка світлих тонів.
Черевний бік, як правило, має незначну дрібну плямистість, що створює «мармурове» забарвлення черева та горла. Від переднього краю морди часто простягається вузька темна смужка, що проходить через ніздрі, очі й далі (часто в неясно виявленому вигляді) через барабанну перетинку. На передніх та задніх кінцівках можуть бути темні поперечні плями-смуги.
Задні кінцівки помірної довжини (коротші, ніж в озерної жаби, але довші, ніж у ставкової) і якщо їх розташувати перпендикулярно поздовжній осі, то надступаковогомілкові (скакові) зчленування звичайно дещо заходять одне за одне. П'ятковий горбок дещо більший (вищий), ніж у Р. ridibundus, але дрібніший (нижчий), ніж у Р. lessonae.
Для самців їстівної жаби, так само як і для більшості безхвостих земноводних, характерний розвиток на перших пальцях передніх кінцівок темно-брунатних шлюбних мозолів. Крім того, вони мають парні резонатори сірого (брудно-білого) кольору.
Мінливість
Матеріали з мінливості зовнішньоморфологічних ознак P. esculentus в Україні обмежені. На Закарпатті розміри постметаморфозних тварин 27,6-100 мм, а середні величини більшості індексів в об'єднаній вибірці статевозрілих тварин (L = 49,03-86,98) коливаються у межах: F./T. = 1,0-1,1, L. c./Lt. c. = 1,1-1,3, L. c./L. o. = 3,9-5,3, L. o./L. tym. = 1,3-1,4, L. c./D. r.-o. = 2,8-3,2, Sp. ip. /D. r.-o. = 0,7-0,9, D. p./D. r.-o. = 2,1-2,4, T./D. r.-o. = 7,3-8,5, L./T. = 2,1-2,3, L./F. + T. = 25,9-39,6. Ці показники дещо відрізняються від ознак P. esculentus з Київської та Черкаської областей.
Порівняння окремих ознак P. esculentus закарпатських тварин з зеленими жабами зі східних ділянок за межами України (Удмуртія і Мордовія, Росія) показало, що останні почасти відрізняються від особин з Закарпаття за ознаками F./T., L./T., T./L. t. ci. і D. p./L. t. ci. Особливо цікавою є мінливість діагностичного індексу D. p. / L. t. ci. у Р. lessonae, P. esculentus та P. ridibundus, який у цих амфібій в Удмуртії практично не перекривається — відповідно 1,6 (1,4-1,8), 2,0 (1,9-2,2) і 2,7 (2,3- 3,2) (у дужках вказані межі мінливості).
Хоча в зелених жаб у сусідньому з Україною центрально-чорноземному регіоні Росії при відмінах середніх значень цієї ознаки (1,7, 2,2 і 2,9), межі його мінливості перекриваються: 1,3-2,1, 1,6-2,7, 2,3-3,7 (P. lessonae, P. esculentus та P. ridibundus відповідно). Приблизно така сама картина відзначаються й для жаб цих таксонів мордовських популяцій: самці — середні значення D. p./L. t. ci. 1,7, 2,2 і 2,6 (межі мінливості: 1,2-2,1, 1,8-2,5, 2,1-3,2), самиці — середні значення 1,7, 2,3, 2,8 (межі мінливості: 1,3-2,0, 2,2-2,4, 2,1-3,2).
Вивчення мінливості зовнішньоморфологічних ознак представників цієї групи у Західній Європі потвердило, що для багатьох з них характерне часткове перекривання з аналогічними показниками батьківських видів (P. ridibundus та P. lessonae).Діагностичний індекс T./L. t. ci. у P. esculentus у Франції коливається у межах 8-9,5 або 9-10,4, у Німеччині 6,5-8,6, у Нідерландах 6-8,5, у Польщі 7-9, в Румунії 7-9,5.
Особливу цікавість викликає питання можливого впливу на морфологію гібридогенних жаб генетичного оточення (популяційної системи). Цей вплив визначається тим, з ким з батьківських таксонів співіснує P. esculentus. З'ясувалося, що виразної кореляції між морфометрією жаб P. esculentus та популяційними системами не існує. Однак P. esculentus одноманітніші за своїми характеристиками порівняно з батьківськими видами, оскільки спадкований клонально геном не дає рекомбінантів. Та це справедливо для однієї гібридної лінії, що ж до різних ліній P. esculentus, то між ними є велика різноманітність.
Частота різного роду аномалій у зовнішній морфології у їстівних жаб, порівняно з батьківськими таксонами достовірно вища, понадто, в них було також зареєстровано два випадки гермафродитизму.
Вивчення особливостей забарвлення їстівних жаб українських популяцій рідше приваблювало увагу батрахологів. Разом з тим є дані про те, що P. esculentus з околиць Києва за кількістю смуг чи плям на кінцівках мають проміжне положення між батьківськими видами.
В описові каріотипу їстівної жаби з України показана його специфічність, яка полягає в гетероморфності 12-ї пари хромосом, представленої мета- та субметацентриком (або, рідше, субмета- та субтелоцентриком). Ця особливість пов'язана з гібридогенним походженням P. esculentus. Про це свідчить вивчення каріотипу амфібій цього комплексу з Німеччини, у якому доведено наявність маркерних хромосом в P. esculentus на основі порівняльної морфометрії хромосом усіх трьох таксонів, що дозволило показати метацентричний характер гомологів 12-ї пари Р. ridibundus, субметацентричний у Р. lessonae та наявність у цій парі одного метацентрика та одного субметацентрика у P. esculentus.
Серед найцікавіших відкриттів, зроблених при вивченні каріотипів зелених жаб слід відмітити знахідки особин з триплоїдним набором (3n = 36; варто ще раз нагадати, що батьківські види у соматичних клітинах мають по 26 хромосом). При цьому в P. esculentus спостерігалася значно нижча частота вторинних перетягів на 11-й (як і на деяких інших) парі гомологів порівняно з Р. ridibundus.
Вивчення каріотипів показало, що усі триплоїдні тварини мають два хромосомні набори ставкової жаби (26 хромосом) і один озерної (13 хромосом). При схрещуванні диплоїдної самиці Р. ridibundus та триплоїдного самця P. esculentus, а також диплоїдної самиці та триплоїдного самця P. esculentus були одержані триплоїдні самці P. esculentus. Ці результати показують, що триплоїдні самці цього таксона продукують сперматозоїди, що містять два геноми ставкової жаби. Та обставина, що в обох комбінаціях були виключно самці, говорить про те, що в гаметах Р. lessonae скоріш за все міститься фактор визначення чоловічої статі.
Цікавість викликає знахідка в околицях Риги тетраплоїдного самця P. esculentus (4n=52). Однак уважний аналіз фотографії метафазної пластинки з 52 елементами показує, що в цьому випадку може йтися про те, що клітина зафіксована не на стадії метафази, а скоріш за все, анафази. У всякому разі, було б бажано ще раз вивчити це питання.
Одне з перших досліджень з використанням спеціального методу забарвлення (С-забарвлення) хромосом показало виразну специфічність каріотипу P. esculentus порівняно з батьківськими видами, яка полягала у наявності маркерних смуг на коротких плечах 11-ї пари хромосом, чого немає в Р. ridibundus. Хромосомному наборові їстівної жаби властивий гетерозиготний стан — така смуга була лише на одному з гомологів цієї пари.
Використання диференціального забарвлення свідчило про те, що в «гібридному» каріотипі усі хромосоми, що походять від озерної жаби, виразно відрізняються темними ділянками (гранулами) або флуоресцентними плямами в районі первинних перетягів (ці особливості не характерні для хромосом Р. lessonae).
Вивчення амфібій цієї групи в Чехії показало поліморфізм за довжиною 1-ї пари хромосом приблизно в половини досліджених озерних та в половини їстівних жаб. Цей факт дозволяє припустити, що причиною цього явища були події в момент утворення статевих клітин (мейоз) — або транслокація, або нерівний кроссинговер у Р. ridibundus, що, таким чином, доводить інтрогресію на хромосомному рівні.
Інший метод, не менш широко використовуваний у вивченні популяційно-аксономічних відносин цієї групи, базується на визначенні розміру генома. Вміст ядерної ДНК в еритроцитах Р. ridibundus на 16 % перевищує Р. lessonae та на 8 % — P. esculentus. Понадто, порівняння вмісту ДНК у соматичних (еритроцити) та статевих (спермії) клітинах дозволило припустити виключення (елімінацію) в гібридогенної P. esculentus батьківського набору хромосом Р. lessonae, що веде до утворення гамет, які містять гени лише озерної жаби.
Спеціальні цитогенетичні методи показали, що в гібридів може відбуватися як «недо-», так і «наделімінація» геномів. З урахуванням цього та передачі лише одного генома від триплоїдних батьків (гаплоїдна сперма гібридних самців) було запропоновано іменувати неменделівський тип спадкування в гібридів «мероклональним» (від грецьк. me/roj— частина), а не гемі- чи напівклональним (від грецьк. ?mi — половина), оскільки один геном від триплоїдного набору менше половини.
Згодом цим самим методом було виявлено спорадичне трапляння самців гібридогенного таксона, які можуть альтернативно передавати наступному поколінню або геном Р. ridibundus, або геном Р. lessonae.
Раніше вважалося, що нежиттєздатність потомства від схрещування батьківських особин P. esculentus зумовлено накопиченням шкідливих мутацій. У зв'язку з цим особливу цікавість викликають одні з останніх результатів вивчення цієї групи в Швейцарії, якими були підтверджені попередні дані про те, що схрещування гібридогенних P. esculentus усе ж може давати в своєму потомстві життєздатних Р. ridibundus., однак це справедливе для гібридогенних жаб однієї популяції.
Якщо ж у схрещуванні братимуть участь батьківські P. esculentus, що походять від батьків з різних регіонів (з різних геміклонів), тобто такі, що фактично мають різні за походженням геноми озерних жаб (різними шкідливими мутаціями), то останні не будуть виявлятися, даючи життєздатних Р. ridibundus.
Спершу повідомлялося, що висунуте теоретичне обґрунтування підтверджено практичними результатами, однак згодом доволі висока життєздатність потомства виявлена й при схрещуваннях P. esculentus з однієї вибірки.
Нещодавно виявлені триплоїдні та тетраплоїдна особини з популяцій P. esculentus Харківської, Луганської та Донецької областей. У численному відношення поліплоїди становили 24 %. При цьому одні триплоїди мали генотип з двох геномів ставкової жаби та одного озерної (54,8 %), інші — з двох геномів озерної та одного ставкової (41,5 %). У поширенні поліплоїди пов'язані з середньою течією р. Сіверський Донець.
Цікаво, що цей показник (кількість ядерної ДНК) у жаб гібридогенного таксона в радіоактивно забруднених після чорнобильської катастрофи регіонах на 4 % менший, ніж в особин з незабруднених районів. На відміну від P. esculentus, у тварин батьківського виду, Р. lessonae, розмір генома не зменшується (порівнювалися ділянки з рівнем радіації 15 g/год., 60 g/год. і 200 g/год.). Особливості організації генома P. esculentus та специфіка популяційно-генетичних процесів у комплексі зелених жаб чимало залежить від того, з ким з батьківських видів співіснує гібридогенна їстівна жаба. Сполучення таксонів зелених жаб, що живуть в одному місці, дістало назву «популяційних систем». Вони позначаються початковими літерами назв таксонів: R — Р. ridibundus, L — Р. lessonae і E — P. esculentus.
Так, українській та румунській частинам дельти Дунаю властивий комплекс (популяційна система) R-E. У популяціях зелених жаб околиць Києва представлені усі таксони (Р. ridibundus, Р. lessonae та P. esculentus), при цьому в одній з них відзначено майже дво- чи трикратне переважання P. esculentus (61 % проти 33 та 19 %).
У Волинській області є популяції цієї групи, що складаються лише з озерних жаб і такі, що характеризуються наявністю усіх трьох таксонів. У Закарпатській області виявлена популяція їстівних жаб представлена лише самицями. Висунуто припущення, що найвірогідніший спосіб розмноження P. esculentus може бути пов'язаний з партеногенезом, що згодом не підтвердилося. Тут слід зазначити, що у світі поки що взагалі немає однозначних доказів партеногенезу серед амфібій.
У цілому можна зазначити, що існує 4 регіональні групи зелених жаб: 1) з пониззя Дунаю, 2) з рівнинного Закарпаття, 3) з більшої частини Правобережного Лісостепу, Прикарпаття, Поділля та 4) з Середнього Подніпров'я. Для кожного з цих регіонів характерні своя специфіка генетичної структури зелених жаб та скерованість генних потоків, а також плоїдність Р. esculentus.
З досліджень зелених жаб цього комплексу за межами України найбільшу цікавість викликають декілька. Так, виявлено, що на північному сході Польщі живуть «чисті» Р. lessonae, «чисті» Р. esculentus та «мішані» Р. esculentus — Р. lessonae (L-E).
Серед зелених жаб Латвії знайдена популяція P. esculentus, представлена практично самими самцями, — Е-система. У південніших регіонах (Угорщина) в заплавних ділянках Дунаю поширені 2 або 3 типи популяційних систем. Перша: L-E-R та L-E (при цьому відзначається дуже низька чисельність самців P. esculentus); друга: L-E у трьох варіантах (пробування самих самиць P. esculentus та Р. lessonae; Р. lessonae співіснує з P. esculentus в однаковому співвідношенні; представлені в основному P. esculentus, що живуть з декількома особинами Р. lessonae); третя: R-E та L-E-R.
Серед зелених жаб Центральної Європи у двох основних типів популяційних систем (L-E і R-E) трапляється декілька варіантів.
| Тип популяційної системи: L-E | Тип популяційної системи: R-E |
|---|---|
| L + 2nE самиці й самці | R + 2nE самці й самиці |
| L + 2nE самиці | R + 2nE самці |
| L + 2nE та 3nE самиці й самці | R + 2nE та 3nE самці й самиці |
| L + 3nE самці | R + 3nE самці |
Ці дані доповнені уточненнями про поширення в Європі 7 типів популяційних систем (послідовність генотипів вказує на їхню відносну частоту): R + L + E; L + E або E + L; E самиці; L + E самці; E + R; R + E самці; E.
У системах «R + E самці» особини чоловічої статі гібридогенного таксона формують гамети з геномом ставкової жаби, хоча в системах «L + E» сперматозоїди P. esculentus містять геном Р. ridibundus.
Останні дослідження жаб цього комплексу на Балканах дозволили виявити тут дві популяції P. esculentus, що були представлені тільки самицями.
Слід підкреслити, що генетичне оточення (тип популяційної системи) справляє певний вплив на мінливість P. esculentus. Однак, як вказувалось раніше, виразної кореляції між морфометрією жаб P. esculentus та популяційними системами не існує. При цьому можна відзначити різну чисельність триплоїдних гібридогенних особин у Центральній Європі: наприклад, в одній з проаналізованих популяцій вони становили 47,5 %, в іншій — 21,9 %.
Самі ж P. esculentus одноманітніші за своїми характеристиками у порівнянні з батьківськими видами, більшість триплоїдних самиць мали генотип LLR. Що ж до триплоїдних самців, то припускається, що у зв'язку зі своєю відносно більшою чисельністю (порівняно з диплоїдними самцями та триплоїдними самицями) вони частіше включаються у схрещування.
У східних районах поширення зелених жаб (в основному територія Росії) показано існування 4-7 різних типів популяційних систем. Викликає цікавість одне з останніх припущень про те, що в популяціях, що розмножуються і які складаються лише з P. esculentus, поряд с гібридними дорослими особинами, щорічно виникають і не-гібриди, але вони гинуть протягом личинкового розвитку. Встановлено, що генотипи гібридних особин представлені трьома варіантами: перший — двома геномами ставкової та одним озерної, другий — одним геномом ставкової та одним озерної, третій — одним геномом ставкової та двома озерної. При цьому майже всі самиці диплоїдні, а самці — триплоїдні. Більшість триплоїдів характеризувалася наявністю двох геномів ставкової жаби та одного озерної.
Інтродукція Р. ridibundus у регіони з аборигенною Р. lessonae згодом, скоріш за все, призводить до її витіснення за допомоги генетичного механізму. Цей механізм являє собою витіснення аборигенних ставкових жаб внаслідок первинної гібридизації озерної та ставкової жаб (потомство представлено їстівною жабою), гібридогенезу, тобто схрещування P. esculentus з Р. ridibundus (потомство представлено озерною жабою) та продукування життєздатного потомства у схрещуваннях P. esculentus.
Спеціальний аналіз шлюбних криків зелених жаб трьох таксонів з колишньої Югославії (з урахуванням температури порівнювали декілька параметрів: інтервал між криками, його тривалість, кількість пульсів за секунду тощо) показав, що їстівні жаби характеризуються проміжним станом цих параметрів порівняно з батьківськими видами Р. ridibundus та Р. lessonae. З іншого боку, основна частина параметрів шлюбних криків Р. esculentus демонструє в них наявність істотної частини модифікацій шлюбних криків Р. ridibundus. Самиці ставкової та їстівної жаб віддають перевагу шлюбним крикам самців Р. lessonae.
Життя у природі



Точні відомості про початок сезону активності їстівних жаб відсутні, однак вони навряд чи сильно відрізняються від цих строків у батьківських видів. Найвірогідніше, вихід їх з місць зимівлі також зумовлений погодними умовами весни щороку й припадає на кінець березня — квітень.
Дещо пізніше починається нерест, який зумовлений специфікою взаємостосунків з батьківськими видами у цій групі та її популяційною структурою. Нерест триває майже до кінця червня — липня.
При дворічному вивченні мішаної популяції з усіх трьох таксонів зелених жаб у Швейцарії, було встановлено, що формування шлюбних пар (та відкладання ікри) супроводжувалося істотним відхиленням від випадкового. Так, парування всередині батьківського виду Р. lessonae було частішим або таким самим, як і в комбінаціях з участю жаб інших таксонів, ніж можна було сподіватися виходячи з їхньої чисельних співвідношень.
В утворенні шлюбних пар часто відзначалися пари самців їстівних жаб з великими самицями Р. lessonae, пари ж між самицями та самцями Р. esculentus якщо й відзначали, то вкрай рідко. Таким чином, існує припущення про те, що репродуктивна ізоляція у групі зелених жаб може почасти забезпечуватися за рахунок міжвидових відмінностей у розмірах тіла.
Відомості про характеристики ікри та личинок Р. esculentus у більшості праць, що передували описові гібридогенного комплексу зелених жаб, також обмежені складнощами в ідентифікації цього таксона і, скоріш за все, можуть стосуватися також пуголовків озерної та/або ставкової жаб. На найбільшу довіру заслуговують праці експериментального характеру, у яких батьки личинок відомі достеменно.
Так, при вивченні потомства від двох самиць їстівної жаби з Польщі з'ясувалося, що відкладена ікра формує три розмірних класи: дрібні ікринки (діаметр 1,09-1,28 мм), середні (1,31-1,56 мм) та великі (1,59-1,89 мм), які в численному відношенні становили 1,1 і 2,1 %, 97,3 і 97,7 %, 0,6 і 1,2 % відповідно. Після хромосомного аналізу личинок, що вивелися з цієї ікри виявилося, що з дрібних ікринок 8 дали пуголовків з диплоїдним набором хромосом 2n=26, а дві — триплоїдів з 3n=39; усе проаналізоване потомство з 16 середньорозмірних ікринок були диплоїдами з 2n = 26, а личинки з великих ікринок — триплоїдами з 3n=39.
Вивчення особливостей міграції дорослих тварин групи (Р. esculentus, Р. lessonae, Р. ridibundus) показало, що частина жаб звичайно мігрує між сусідніми ставками, що їх вони займають для нересту. Так, з 2735 помічених жаб 3-10 % мігрували з однієї водойми до іншої протягом сезону активності і приблизно така ж сама кількість (2-12 %) — між сезонами.
Їстівні жаби можуть зимувати як у воді (при їх спільному пробуванні з Р. ridibundus), так і на суші (у випадку співіснування з Р. lessonae). Особливу цікавість викликає вивчення специфіки зимівлі їстівних жаб на озері Neusiedl в Австрії. Так, тут було встановлено, що Р. esculentus у виборі місць зимівлі віддавали перевагу озеру чи довколишним канавам, зроблені незвичайні знахідки разом з Р. lessonae на зимівлі декількох особин Р. esculentus за 15 км від місць мічення.
Спеціальне дослідження живлення цього виду не проводилося, однак можна припустити, що за складом здобичі Р. esculentus навряд чи відрізняється від представників інших таксонів цього комплексу. Відомості про специфіку реакції на дію антропогенного фактора Р. esculentus порівняно з іншими представниками цього комплексу вкрай обмежені. Серед схожих досліджень слід вказати на працю з вивчення впливу двох чинників (конкуренції та пересихання водойм) на пуголовків ставкової та їстівної жаб (Semlitsch, 1991). Показано, що личинки перших швидше проходять метаморфоз в умовах постійного рівня водойм. Якщо ж водойми характеризуються тенденцією до пересихання, то личинки Р. esculentus дістають у такому разі перевагу. Перевага пуголовків їстівних жаб також виявлялася в умовах їхнього розвитку при підвищеній щільності.
Охорона
Вид занесено до Додатку III «Конвенції з охорони дикої флори і фауни та природного середовища існування в Європі» (категорія «види, що підлягають охороні») та до «Червоної книги хребетних Міжнародного союзу охорони природи (МСОП)».
Посилання
Писанець Є. Земноводні України (посібник для визначення амфібій України та суміжних країн). — Київ : Вид-во Раєвського, 2007. — 197 с.