Міксоміцети
Міксоміце́ти (від грец. μύξα — слиз і μύκητος — гриб), слизовики́ (Mycetozoa) — група найпростіших клади Amoebozoa, що за зовнішнім виглядом, наявністю плодових тіл, та способом життя нагадують гриби, проте відрізняються клітинною організацією, особливостями розмноження та життєвого циклу і не споріднені із ними філогенетично[1]. Первинно-гетеротрофні тубулокристати.
? Міксоміцети | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
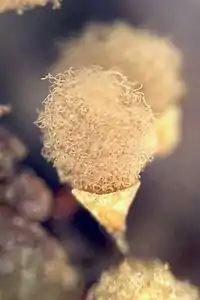
 Плодові тіла Badhamia utricularis | ||||||||||||||
| Біологічна класифікація | ||||||||||||||
| ||||||||||||||
Посилання | ||||||||||||||
| ||||||||||||||
Класифікація
Слизовики відкрили в середині 18 століття (Battara, 1755; Haller, 1768) і класифікували як гриби на основі таких спільних рис, як розмноження і поширення спорами та утворення плодових тіл[2]. 1859 року Антуан де Барі вилучив міксоміцети із царства грибів через те, що вони мають амебоподібні клітини без клітинних стінок і живляться шляхом фагоцитозу[3]. Він запропонував для них назву Mycetozoa (гриби-тварини), що відображала подвійну природу цих організмів, і відніс до тварин, вказавши на спорідненість із саркодовими (Sarcodina)[2]. Місце міксоміцетів у системі живого довго залишалось невизначеним, у різних класифікаціях вони належали як до тварин, так і до грибів (наприклад, у класифікації Роберта Віттекера), а інколи деяких з них відносили навіть до царства рослини[4].
Наприкінці 1960-х років усталилася морфологічна класифікація Джорджа Мартіна та Константиноса Алексопулоса[5], заснована на всебічному аналізі будови слизовиків: наявності чи відсутності в плодових тілах ниток, їх розгалуженням, складу мінеральних відкладів (оксалатів, карбонатів, фосфатів кальцію). За цією класифікацією ендоспорові («справжні») міксоміцети поділяли на пять порядків[6]:
- ехіностелієві
- ліцеєві
- трихієві
- стемонітові
- фізарові.
Раніше до слизовиків включали також акразієвих, лабіринтуломікотових і плазмодіофорових (Plasmodiophorida), проте структурні (а пізніше — і молекулярні) дані засвідчили їх неспорідненість із міксоміцетами[4][2].
На початку 2000-х стало зрозуміло, що міксоміцети філогенетично не споріднені зі справжніми грибами, а наявність плодових тіл в обидвох групах організмів є прикладом конвергентної еволюції. На основі даних молекулярних досліджень, а також того факту, що слизовики мають мітохондрії із трубчастими кристами, їх включили до клади Amoebozoa[1][7].
На початку XXI сторіччя у складі Mycetozoa виділяли три основні групи, що відрізняються особливостями життєвого циклу[4]:
- міксомікоти (Myxogastria, справжні або плазмодіальні слизовики) — мають життєву стадію плазмодію, який є однією великою багатоядерною клітиною (до мільйонів ядер);
- диктіостеліди (Dictyostelia/Dictyostelea, клітинні слизовики, Dictyostelium discoideum) — амебоїдні одноклітинні організми, що за несприятливих умов можуть збиратись у багатоклітинні псевдоплазмодії
- протостеліди (Protostelia) — мікроскопічні амеби, що утворюють одиночні спори на ніжках. Філогенетичне дослідження, яке було опубліковано 2009 року, показало, що протостеліди взагалі не утворюють монофілетичної групи, а складаються щонайменше з шести різних ліній, які розкидано між доволі віддаленими гілками супергрупи амебозоїв (Amoebozoa)[8]. До середини 2010 років клас протостелід остаточно розпався, а його колишнім представникам знайшли відповідні місця на філогенетичному дереві амебозоїв. Порядок Protosporangiida (до якого входять роди протоспорангіум, кластостеліум і цераціомікса) виявся близькоспорідненим до міксоміцетів[9][6].
Особливості будови
Місомікотові
Плазмодіальні або справжні слизовики на вегетативній стадії представлені багатоядерним плазмодієм, який утворюється внаслідок серії послідовних мітозів без цитокінезу. Вони часто мають яскраве забарвлення, переважно жовте або помаранчеве. Цитоплазма всередині плазмодію постійно рухається і пульсує, це сприяє транспорту поживних речовин та кисню.
Плазмодій може рухатися у бік вологого місця та назустріч потоку води. На основі цього існує метод виманити міксоміцета, наприклад, з трухлявої деревини. Для цього в глибину старого пенька вкладають смужку скла, накриту папером, кінець якого поміщають у ємність з водою. Плазмодії будуть рухатися на скло, приманені рухом води [10]. Після періоду росту міксоміцет рухається у бік світла. Вважають, що світло потрібне для утворення спорангій, позаяк розповсюджувати спори краще на поверхні[11].
Плазмодій міксомікотових має досить незвичний вигляд, через що інколи виникають непорозуміння. Наприклад, 1973 року житель Далласу виявив у себе на подвір'ї пульсуючу червону масу діаметром півметра, про що сповістив пресу. Служби новин стали повідомляти про відкриття «нової форми життя», яка багато кому нагадала про фільм 1958 року Крапля. Згодом біологи ідентифікували масу як плазмодіальний слизовик[12].
Диктіостеліди
Трофічна стадія розвитку диктіостелід представлена поодинокими амебоподібними клітинами, які за несприятливих умов можуть збиратись у псевдоплазмодій, зовнішньо схожий на плазмодій міксомікотових, проте відмінний від нього багатоклітинністю[2].
Живлення
Слизовики переважно сапротрофи, живуть у вологому ґрунті, на гнилій деревині та листі. Живляться дрібними грибами, бактеріями, органічними рештками шляхом фагоцитозу[7].
Життєвий цикл
Плазмодіальні слизовики
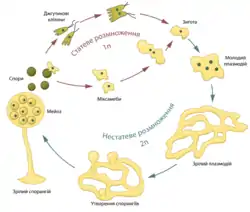
Вегетативна стадія міксомікотових слизовиків є диплоїдною. Коли плазмодій дозріває або потрапляє в умови нестачі їжі або/і вологи, він перетворюється на плодові тіла (спорангії), в яких після дозрівання шляхом мейозу утворюються спори, вкриті целюлозними оболонками. На електронних мікрографіях спор Didymuim nigripes видео два шари: внутрішній целюлозний, зовнішній хітиновий, а у Physarum gyrosum спостерігають навіть три[13]. Ці оболонки дозволяють спорам зберігатися тривалий час, поки вони не потраплять у сприятливі умови.
Для розповсюдження спор у спорангії багатьох міксомікотових наявні особливі ниткоподібні волокна, які утворюють спіральну мережу, так званий капіліцій. У незрілій спорангії вони стиснуті, проте після розривання оболонки розправляються, розпушуючи спори та викидаючи їх назовні[11].
За наявності води спори проростають, і з них виходять безджгутикові клітини (міксамеби) або джгутикові (зооспори), які можуть перетворюватись одні на одних. Ці клітини гаплоїдні, вони деякий час активно живляться, після чого попарно зливаються (міксамеби — з міксамебами, джгутикові клітини — із джгутиковими клітинами), внаслідок чого утворюються диплоїдна зигота. Зигота живиться й росте, її ядро багаторазово ділиться шляхом мітозу, при цьому не відбувається цитокінезу; таким чином з'являється молодий плазмодій[1][7]. Новостворений організм рухається у глибину середовища, у якому він живе (пеньки, опале листя), де росте до нового спороношення. Спостерігається об'єднання кількох маленьких плазмоїдів у один організм. Цей механізм раніше вважали процесом народження слизовиків, проте зараз доведено можливість існування в одному плазмодії ядер різного походження (гетерокаріоз). Наприклад у Didymium nigripes знайдено ядра з різною кількістю ДНК[13].
При несприятливих умовах плазмодій може покриватися твердіючою речовиною (склероцій). У такому стані він може перечекати посуху. Зафіксовано випадок перетворення міксоміцета (зі склероція на плазмодій), що пролежав у гербарії 20 років[11].
Клітинні слизовики
На відміну від плазмодіальних слизовиків у клітинних вегетативна стадія — одноклітинні міксамеби — гаплоїдна. За певних умов міксамеби можуть зливатись між собою, формуючи зиготу, яка покривається товстою оболонкою, після чого проходить мейотичний та кілька мітотичних поділів. Утворені гаплоїдні міксамеби вивільняються у середовище внаслідок розриву оболонки[7]. У випадку нестачі їжі диктіостеліди розмножуються нестатево. Клітини виділяють цАМФ, який приваблює інші міксамеби, що в свою чергу також починають синтезувати цю сигнальну сполуку. Таким чином вони агрегують і утворюють багатоклітинний псевдоплазмодій, що деякий час рухається, залишаючи за собою слизовий слід. Згодом розпочинається диференціація клітин, частина з них формує стерильну ніжку плодового тіла (сорокарпа, відрізняється від спорангія тим, що спори формуються екзогенно (зовнішньо)[2]), інша частина утворює спори. Спори гаплоїдні й оточені захисними оболонками, проростають за сприятливих умов[1].
Інтелект
В одному з дослідів вчені змінювали температуру Physarum polycephalum з холодної на теплу з інтервалом 60 хвилин і помітили, що слизовик готується до зміни «погоди», коли наближається час. Цей дослід повторювали з різними інтервалами часу (30 і 90 хвилин), і у всіх випадках слизовик «запам'ятовував», коли має змінитися температура.
Відомий експеримент із проходженням лабіринту був проведений з Physarum polycephalum. Під час руху він залишає після себе сліди слизу. Ці краплі слизу організм використовує як орієнтири в просторі, і якщо не може кудись потрапити, то по цих стежках може легко повернутися назад.[14]
Слизовики як модельні об'єкти
Багато видів слизовиків легко культивувати, їхній життєвий цикл недовгий і минає за кілька днів, це зробило їх зручним модельним об'єктом. Зокрема, їх використали при дослідженні цитоскелету, функціонування кальцій-кальмодулінового комплексу, еволюції багатоклітинності та статевого процесу, ієрархії ядер в багатоклітинних структурах та багатьох інших процесів.
Dictyostelium discoideum використовували для вивчення сигнальної молекули цАМФ (яку інколи називають акразином, через те, що D. discoideum раніше відносили до акразієвих), яку клітини цього слизовика використовують для агрегації[2]. Також дослідження цього виду слизовиків дозволили пролити світло на деякі питання еволюції багатоклітинності. Життєвий цикл диктіостелуму передбачає «самопожертву» частини міксамеб, через те, що при формуванні плодового тіла (сорокарпа) репродуктивний потенціал мають тільки ті клітини, що диференціюються в спори; ті ж, що потрапляють до складу стерильної ніжки, виконують тільки допоміжну функцію і гинуть, так і не передавши спадкову інформацію нащадкам. У деяких представників спостерігається мутація, внаслідок якої вони стають «шахраями» і ніколи не беруть участі в утворенні ніжки. Така особливість, очевидно, надає клітинам значної репродуктивної переваги, проте не поширюється в популяції. Це пов'язане з тим, що «чесні» міксамеби можуть розпізнати мутантів за відсутністю певного білка на поверхні і не агрегувати з ними. Схожі системи розпізнавання могли бути важливими для еволюції багатоклітинних еукаріот[7].
Джерела
- Prescott L.M. (2002). Microbiology (вид. 5th). McGraw−Hill. ISBN 0-07-282905-2.
- Глущенко В.И., Леонтьев Д.В., Акулов А.Ю. Слизевики: учебное пособие для ВУЗов. — Харьков : ХНУ, 2002. — 135 с. — ISBN 966-623-107-7.
- Леонтьєв Д.В., Акулов О.Ю. Загальна мікологія: Підручник для вищих навчальних закладів. — Харків : Основа, 2007. — 228 с. — ISBN 978-966-495-040-1.
- Baldauf SL, Doolittle WF. (1997). Origin and evolution of the slime molds (Mycetozoa). Proc Natl Acad Sci USA 94: 12007–12. PMID 9342353.
- G. W. Martin, C. J. Alexopoulos (1969). Myxomycetes. OCLC 652011640.
- Дмитрий Леонтьев (10.04.2019). Предложена первая филогенетическая классификация миксомицетов. «Элементы»: Новости науки. Процитовано 19 жовтня 2020.(рос.)
- Campbell NA, Reece JB (2008). Biology (вид. 8th). Benjamin Cammings. ISBN 978-0321543257.
- Lora L. Shadwick, Frederick W. Spiegel, John D. L. Shadwick, Matthew W. Brown, Jeffrey D. Silberman (25 серпня 2009). Eumycetozoa = Amoebozoa?: SSUrDNA Phylogeny of Protosteloid Slime Molds and Its Significance for the Amoebozoan Supergroup. doi:10.1371/journal.pone.0006754.
- Silar, 2016, с. 238.
- Жизнь растений. Том 2. Грибы, 1976, с. 7.
- Жизнь растений. Том 2. Грибы, 1976, с. 8.
- Tortora GJ, Funke BR, Case CL (2009). Microbiology. An Introduction (вид. 10th). Benjamin Cummings. ISBN 978-0321582034.
- Жизнь растений. Том 2. Грибы, 1976, с. 9.
- Jabr, Ferris. How brainless slime molds redefine intelligence. Nature. doi:10.1038/nature.2012.11811. Процитовано 5 грудня 2016.
Посилання
- Грибы // Жизнь растений : [рос.] : в 6-ти т. / Гл. ред. чл. -кор. АН СССР, проф. Ал. А. Федоров. — Просвещение. — М., 1976. — Т. 2. — 479 с. : ил.
- Костіков І. Ю., Джаган В. В., Демченко Е. М., Бойко О. А., Бойко В. Р., Романенко П. О. Ботаніка. Водорості та гриби. — Київ, 2004.
- Кривомаз Т.І. Міксоміцети України[недоступне посилання з липня 2019] — 2010 — Переглянуто 1 квітня 2012 року
- Глущенко В.И., Леонтьев Д.В., Акулов А.Ю. Слизевики: учебное пособие для ВУЗов. — Харьков : ХНУ, 2002. — 135 с. — ISBN 966-623-107-7.(рос.)
- Змитрович, И.В. Рецензії : [… книга претендует скорее на статус учебника, нежели методического пособия. Однако возможность использования рецензируемой работы в этом статусе вызывает определенные опасения … Налицо дидактическая и терминологическая небрежность авторов … Проводимый авторами исторический экскурс изобилует неточностями и ложной информацией.] : [рос.] // Український ботанічний журнал. — Т. 61, вып. № 4. — С. 109—112. — ISSN 0372-4123. — Рец. на кн.: Глущенко В.И., Леонтьев Д.В., Акулов А.Ю. Слизевики. Учебное пособие. — Харьков : ХНУ, 2002. — 135 с.
- Stephenson SL (2000). Myxomycetes: A Handbook Of Slime Molds. Timber Press. ISBN 978-0881924398.
- D. V. Leontyev, M. Schnittler, S. Stephenson, Y. K. Novozhilov, O. N. Shchepin (2019). Towards a phylogenetic classification of the Myxomycetes. Phytotaxa 399 (№3): 209–238. doi:10.11646/phytotaxa.399.3.5.
- Silar, Philippe (2016). Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes. HAL archives-ouvertes. с. 462. ISBN 978-2-9555841-0-1.(фр.)
- Відео Агрегація Dyctiostelium
- Відео Physarum sp.
