Регуляторна Т-клітина
Регуляторні Т-клітини (Treg [ˈtiːrɛɡ]), раніше відомі як супресорні Т-клітини, належать до субпопуляції Т-клітин, вони модулюють імунну систему, підтримують толерантність до власних антигенів, і запобігають аутоімунним захворюванням. Treg - імуносупресивні клітини й зазвичай або пригнічують вже наявні ефекторні Т-клітини, або блокують їх появу [1] Treg експресують біомаркери CD4, FOXP3 і CD25 і, як вважають, походять з тієї ж лінії, що й наївні CD4+ клітини.[2] Оскільки ефекторні Т-клітини також експресують CD4 і CD25, Treg дуже важко надійно відрізнити від ефекторної CD4+ клітини, що ускладнює їх вивчення. Нещодавні дослідження показали, що цитокін трансформуючий фактор росту бета (TGF-β) необхідний для диференціації Treg від наївних CD4+ клітин і має важливе значення для підтримки гомеостазу Treg.[3]
На основі мишачих моделей було припущено, що модуляція Treg може лікувати аутоімунні захворювання та злоякісні пухлини, а також може сприяти трансплантації органів [4] та загоєнню ран.[5] Значущість Treg при злоякісних пухлинах складна. Як правило, їх кількість підвищуються у людей із злоякісними пухлинами, і, здається, вони мігрують до місць росту пухлин. Дослідження як на людях, так і на тваринних моделях показали, що велика кількість Treg у мікросередовищі пухлини свідчить про поганий прогноз хвороби. Вважається, що Treg пригнічують протипухлинний імунітет, таким чином перешкоджаючи вродженій здатності організму контролювати ріст ракових клітин.[6] Останні дослідження в галузі імунотерапії присвячені тому, як регуляція Т-клітин може бути використана для лікування раку. [7]
Популяції
Treg є компонентом імунної системи, який пригнічує імунні реакції інших клітин. Це важлива «самоперевірка», вбудована в імунну систему, щоб запобігти надмірним реакціям. Treg існують у багатьох формах, найбільш добре вивченими є ті, які експресують CD4, CD25 і FOXP3 (CD4+ CD25+ Treg). Ці Treg відрізняються від Т-хелперів.[8] Іншою підгрупою регуляторних Т-клітин є Treg17.[9] Treg беруть участь у припиненні імунних реакцій після того, як вторгнення патогенів було успішно подолано, а також у запобіганні аутоімунітету.[10]
CD4+ FOXP3+ CD25(high) Treg були названі «природними» Treg [11] щоб відрізнити їх від «супресорних» популяцій Т-клітин, які генеруються in vitro. Інші популяції Treg включають Tr1, Th3, CD8+ CD28− і Qa-1 обмежені T-клітини. Внесок цих популяцій у аутотолерантність та імунний гомеостаз досліджений менше. FOXP3 можна використовувати як хороший маркер для мишачих CD4+ CD25+ T-клітин, хоча останні дослідження також надали докази експресії FOXP3 у CD4+ CD25− T-клітин. У людей FOXP3 також експресується у нещодавно активованих звичайних T-клітин і, таким чином, не є специфічними для людських Treg.[12]
Розвиток
Усі Т-клітини походять від клітин-попередників у кістковому мозку, які пов'язані зі своєю лінією в тимусі. Усі Т-клітини починаються як CD4- CD8- TCR-клітини на стадії DN (double-negative, подвійно негативні), коли в окремій клітині йде перебудова генів Т-клітинних рецепторів, що формує унікальну функціональну молекулу, яку ці клітини, у свою чергу, "тестують" в корі тимуса для забезпечення мінімального рівня взаємодії з власними молекулами ГКГС. Якщо Т-клітини отримують такі сигнали, вони проліферують і експресують як CD4, так і CD8, стаючи "подвійно позитивними" (DP) клітинами. Виділення Treg відбувається з радіорезистентних гемопоетичних клітин, що експресують МНС класу II, у мозковій речовині або у тільцях Гаселя в тимусі. На стадії DP (подвійно позитивні) вони відбираються шляхом взаємодії з клітинами тимуса, починають транскрипцію Foxp3 і стають Treg, хоча вони можуть не почати експресувати Foxp3 до однопозитивної стадії, у який момент вони стають функціональними. Експресія TCR у Treg не обмежена, як у NKT або γδ Т-клітин. Treg мають більшу різноманітність TCR, ніж ефекторні Т-клітини, і ці рецептори більше подібні до власних пептидів.
Процес селекції Treg визначається спорідненістю взаємодії з комплексом "аутопептид-ГКГС". Процес відбору клітин для отримання статусу Treg схожий на казку "Три ведмедя", тобто сигнал має бути не надто високим, не надто низьким, але "якраз".[13] Т-клітина, яка отримує дуже сильні сигнали, зазнає апоптозу; клітина, яка отримує слабкий сигнал, виживе і стане ефекторною клітиною. Якщо ж Т-клітина отримує проміжний сигнал, то вона стане регуляторною Т-клітиною. Через стохастичну природу процесу активації усі популяції Т-клітин із певним TCR отримують суміш ефекторних та регуляторних T-клітин – їх відносна пропорція визначаються спорідненістю Т-клітини до комплексу "аутопептид-MHC". Навіть у мишачих моделях мишей з TCR-трансгенними клітинами, відібраними на стромі, що секретує специфічний антиген, делеція або конверсія не є повною.
Генерація Foxp3+ Treg у тимусі затримується на кілька днів порівняно з ефекторними Т-клітинами і не досягає дорослого рівня ні в тимусі, ні на периферії до терміну приблизно три тижні після пологів. Клітини Treg потребують костимуляції CD28, а експресія B7.2 значною мірою обмежена мозковою речовиною, розвиток якої, здається, іде паралельно розвитку клітин Foxp3+. Було висловлено припущення, що вони пов’зані між собою, але остаточного зв'язку між процесами поки не виявлено. TGF-β не потрібен для функціональності Treg у тимусі, оскільки Treg вилочкової залози у мишей, нечутливих до TGF-β (миші TGFβRII-DN) є функціональними.
Рециркуляція в тимусі
Помічено, що деякі FOXP3+ Treg рециркулюють назад у тимус. Такі Treg в основному були присутні у мозковій речовині тимуса, який є основним місцем диференціювання Treg.[14] Наявність цих клітин у тимусі або додавання в культуру тканини тимуса плода пригнічує розвиток нових Treg на 34 – 60%,[14] але на CD4+CD25− конвенційні Т-клітини це не впливає. Це означає, що рециркуляційні Treg гальмують тільки de novo розвиток Treg. Молекулярний механізм цього процесу працює завдяки здатності Treg адсорбувати IL-2 з мікросередовища, таким чином, вони знатні індукувати апоптоз інших Т-клітин, які потребують IL-2 як основного фактора росту.[15] Рециркулюючі Treg клітини в тимусі експресують високу кількість α-ланцюга рецептора IL-2 (CD25), що кодується геном Il2ra, цей ланцюк "збирає" IL-2 з мозкової речовини тимуса і знижує його концентрацію. Новостворені FOXP3+ Treg тимусу мають не такий високий рівень експресії Il2ra.[14] IL-2 є цитокіном, необхідним для розвитку Treg в тимусі. Він важливий для проліферації та виживання Т-клітин, але у разі його дефіциту він може бути заміщений IL-15. Проте розвиток Treg залежить від IL-2.[16] У людей була виявлена популяція CD31- Treg тимуса.[14] CD31 можна використовувати як маркер новоутворених Treg так же, як і інших Т-лімфоцитів. Зрілі та периферичні Treg зменшують його експресію.[17] Отже, можливо, що цей регуляторний механізм розвиток Treg у тимусі також є функціональним у людини.
Ймовірно, існує також позитивна регуляція розвитку Treg у тимусі, викликана рециркуляцією Treg клітини в тимус. Виявлено популяцію CD24 (низька експресія) FOXP3+ клітин у тимусі з підвищеною експресією IL-1R2 (Il1r2) порівняно з периферичними Treg.[18] [19] Висока концентрація IL-1β, викликана запаленням, зменшує розвиток de novo Treg в тимусі.[19] Наявність в тимусі рециркуляційних Treg з високою експресією IL1R2 під час запальних станів допомагають поглинати IL-1β і знижувати його концентрацію в мікросередовищі мозкової речовини, таким чином вони сприяють розвитку de novo Treg.[19] Зв'язування IL-1β з IL1R2 на поверхні Treg не спричиняє передачі сигналу, оскільки немає внутрішньоклітинного (TIR) рецепторного домену Toll інтерлейкіну-1, який зазвичай присутній у клітинах вродженого імунітету. [20]
Функція
Імунна система повинна вміти розрізняти власні та чужі структури. Коли поділ «своє/не своє» зазнає невдачі, імунна система руйнує клітини та тканини власного організму, що в результаті призводить до аутоімунного захворювання. Регуляторні Т-клітини активно пригнічують активацію імунної системи та запобігають патологічній аутореактивності, тобто аутоімунному захворюванню. Про критичну роль регуляторних Т-клітин в імунній системі свідчить тяжкий аутоімунний синдром, який є результатом генетичного дефіциту регуляторних Т-клітин (синдром IPEX – див. нижче).
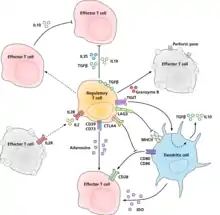
Молекулярний механізм, за допомогою якого Treg здійснюють свою супресорну/регуляторну активність, остаточно не описаний і є предметом інтенсивних досліджень. Експерименти in vitro надали неоднозначні результати щодо необхідності міжклітинного контакту з клітиною, що пригнічується. Нижче наведено деякі із запропонованих механізмів пригнічення імунітету:
- Treg виробляють ряд гальмівних цитокінів. До них належать TGF-β, [21] інтерлейкін 35, [22] та інтерлейкін 10.[23] Також виявляється, що Treg можуть спонукати інші типи клітин експресувати інтерлейкін-10.[24]
- Treg можуть продукувати гранзим В, який, у свою чергу, може індукувати апоптоз ефекторних клітин. Повідомляється, що Treg мишей з дефіцитом гранзиму B є менш ефективними супресорами активації ефекторних Т-клітин. [25]
- Зворотна передача сигналів шляхом прямої взаємодії з дендритними клітинами та індукції імуносупресивної індолеамін-2,3-діоксигенази.[26]
- Передача сигналів через ектоферменти CD39 і CD73 з виробленням імуносупресивного аденозину . [27] [28]
- Завдяки прямій взаємодії з дендритними клітинами LAG3 і TIGIT.[29] [30] Існує літературний огляд взаємодій Treg з дендритними клітинами, що описує відмінність між механізмами, описаними для клітин людини та клітин миші.[31]
- Інший механізм управління - через механізм зворотного зв'язку IL-2. Т-клітини, активовані антигеном, виробляють IL-2, який потім діє на рецептори IL-2 на Treg, попереджаючи їх про те, що в цій ділянці спостерігається висока активність Т-клітин, а Treg, в свою чергу, забезпечують їх супресію. Цей негативний зворотних зв’язок гарантує відсутність надмірної реакції Т-клітин. Якщо інфекція присутня, інші фактори запалення зменшують супресію Т-клітин. Порушення зв'язку призводить до гіперреактивності, регуляція може змінювати силу імунної відповіді.[32] Схожа гіпотеза щодо ролі інтерлейкіну 2 полягає в тому, що активовані Treg захоплюють інтерлейкін 2 настільки активно, що позбавляють ефекторних Т-клітин його достатньої кількості й вони уникають апоптозу.[15]
- Основний механізм супресії полягає в запобіганні костимуляції через CD28 на ефекторних Т-клітинах під дією молекули CTLA-4 .[33]
Індуковані регуляторні Т-клітини
Індуковані регуляторні Т-клітини - iTreg (CD4+ CD25+ FOXP3+ ) є супресивними клітинами, які беруть участь у формуванні імунологічної толерантності. Було показано, що вони пригнічують проліферацію Т-клітин і експериментальні аутоімунні захворювання. До них належать T-хелпери 17. iTreg розвиваються із звичайних зрілих CD4+ Т-клітин поза тимусом, це є визначальною відмінністю між природними Treg (nTreg) та iTreg. Хоча iTreg і nTreg мають подібну функцію, нещодавно було показано, що iTreg є «важливою ненадлишковою регуляторною підгрупою, яка доповнює nTreg, частково за рахунок розширення різноманітності TCR в рамках регуляторних реакцій».[34] Гостре виснаження пулу iTreg у мишачах моделях призводило до запалення та втрати ваги. Конкретний внесок nTreg та iTreg у підтримку толерантності невідомий, але обидва є важливими. Між nTreg та iTreg спостерігалися епігенетичні відмінності, причому перші мають більш стабільну експресію FOXP3 і більш поширене деметилювання.
Середовище тонкого кишечника з високим вмістом вітаміну А є місцем, де виробляється ретиноева кислота.[35] Ретиноева кислота і TGF-бета, що виробляються дендритними клітинами в цій ділянці, сигналізують про продукцію Treg.[35] Вітамін А і TGF-бета сприяють диференціації Т-клітин у Treg, а не у Th17, навіть у присутності IL-6.[36] [37] Кишкове середовище з TGF-бета і ретиноєвою кислотою може призвести до появи індукованих Treg,[38] деякі з яких експресують лектиноподібний рецептор CD161 і спеціалізуються на підтримці цілісності бар’єру шляхом прискорення загоєння ран.[39] Після введення антигену в кишечнику від наївних Т-клітин диференціюються Treg.[40] Нещодавно було показано, що людські Treg можуть бути індуковані як з наївних, так і з попередньо фіксованих клітин Th1 і Th17[41] через дію міметиків TGF-β, отриманих від патогена Heligmosomoides polygyrus, який називається Hp-TGM (H. polygyrus TGF-β міметик).[42] [43] Hp-TGM може індукувати мишачі FOXP3, який експресують Treg, які були стабільними за наявності запалення in vivo. [44] FOXP3+ Treg людини, індуковані Hp-TGM, були стабільними за наявності запалення та мали підвищені рівні CD25, CTLA4 та знижене метилювання в FOXP3 ділянці гена порівняно з TGF-β-індукованим Treg. [41]
Захворювання
Важливим питанням в області імунології є те, як модулюється імуносупресивна активність Treg під час постійної імунної відповіді. Хоча імуносупресивна функція Treg запобігає розвитку аутоімунного захворювання, вона не бажана під час імунних відповідей на інфекції. Сучасні гіпотези припускають, що при зустрічі з патогенами активність Treg може бути знижена, прямо чи опосередковано, іншими клітинами, щоб полегшити ліквідацію інфекції. Експериментальні дані на мишачих моделях свідчать про те, що деякі патогени, можливо, еволюціонували, щоб маніпулювати Treg, для викликання імуносупресії і таким чином вони збільшують шанси на власне виживання. Наприклад, повідомляється, що активність регуляторних Т-клітин підвищується при деяких інфекціях, таких як ретровірусні (найвідомішою з яких є ВІЛ), мікобактеріальні (наприклад, туберкульоз[45]) та різних паразитарних інвазіях, включаючи лейшманіоз і малярію .
Treg відіграють важливу роль під час ВІЛ-інфекції. Вони пригнічують імунну систему, таким чином обмежуючи її відповідь та зменшуючи запалення, одночасно порушуючи виведення вірусу за допомогою клітинно-опосередкованої імунної відповіді та збільшуючи резервуар інфекції, переводячи CD4+ Т-клітини (інфіковані тощо) до стану спокою. Крім того, Treg клітини можуть бути самі інфіковані ВІЛ, що збільшує розмір резервуара вірусу. Через це Treg клітини досліджуються в якості мішені при лікуванні ВІЛ.[46] Деякі стратегії пригнічення Treg були перевірені на нелюдиноподібних приматах, інфікованих SIV і було показано, що вони викликають реактивацію вірусу та посилюють специфічні для SIV CD8+ Т-клітинні реакції.[47]
Treg відіграють велику роль у патології вісцерального лейшманіозу та у запобіганні надмірного запалення у пацієнтів, які вилікувалися від вісцерального лейшманіозу.
CD4+ Treg часто асоціюються з солідними пухлинами як у людей, так і у мишачих моделях. Збільшення кількості Treg при раку молочної залози, колоректальному раку та раку яєчників асоціюється з гіршим прогнозом.[48]
CD70+ В-клітини неходжкінської лімфоми індукують експресію FOXP3 і регуляторну функцію у пухлинних CD4+ CD25- T-клітинах.[49]
Є деякі докази того, що дисфункційні Treg можуть викликати нейрозапалення при боковому аміотрофічному склерозі через нижчу експресію FOXP3.[50] Досліджується метод ex vivo розмноження Treg для подальшої аутологічної трансплантації, після того як були отримані багатообіцяючі результати в клінічному випробуванні фази I.[51]
В той час як було показано, що під час здорової вагітності кількість Treg збільшується внаслідок поліклонального розмноження як системно, так і локально, щоб захистити плід від імунної реакції материнського організму (явище, яке називається материнською імунною толерантністю), є докази того, що це поліклональне розмноження порушується у вагітних з прееклампсією та їх нащадків.[52] Дослідження показують, що зниження продукції та розвитку Treg під час прееклампсії може погіршити материнську імунну толерантність, що призведе до надлишкової імунної відповіді, характерної для прееклампсії.[53]
Злоякісні пухлини
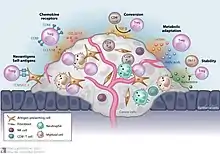
Більшість пухлин викликають імунну відповідь в організмі хазяїна, яка опосередковується пухлинними антигенами, які відрізняють пухлину від інших (непухлинних) клітин. Це призводить до того, що в мікросередовищі пухлини виявляється велика кількість лімфоцитів, що інфільтрують пухлину (TIL).[54] Хоча це не зовсім зрозуміло, вважається, що ці лімфоцити націлені на пухлинні клітини і, отже, уповільнюють або припиняють розвиток пухлини. Однак цей процес є складним, оскільки в мікрооточення пухлини транспортуються переважно Treg. Тоді як зазвичай Treg складають лише близько 4% CD4+ T-клітин, вони можуть становити до 20–30% загальної популяції CD4+-клітин мікросередовища пухлини.[55]
Хоча спочатку вважалося, що високий рівень TIL є важливим для визначення імунної відповіді проти раку, зараз широко визнано, що співвідношення Treg до ефекторних T-клітин мікрооточення пухлин є визначальним фактором успіху імунної відповіді проти раку. Висока кількість Treg у мікросередовищі пухлини пов’язана з поганим прогнозом при багатьох видах раку [56], таких як рак яєчників, рак молочної залози, рак нирок і рак підшлункової залози.[55] Це свідчить про те, що Treg пригнічують ефекторні Т-клітини і перешкоджають імунній відповіді організму проти пухлин. Однак при деяких типах злоякісних пухлин все навпаки, і висока кількість Treg асоціюється з позитивним прогнозом. Ця тенденція спостерігається при таких ракових захворюваннях, як колоректальна карцинома та фолікулярна лімфома. Це може бути пов’язано зі здатністю Treg пригнічувати загальне запалення, яке, як відомо, викликає проліферацію пухлинних клітин і метастазування.[55] Ці протилежні ефекти вказують на те, що роль Treg у розвитку раку сильно залежить як від типу, так і від локалізації пухлини.
Хоча досі не зовсім зрозуміло, як Treg переважно транспортуються в мікрооточення пухлини, вважають, що їх хемотаксис зумовлений хемокінами, що продукуються пухлиною. Інфільтрації Treg у мікросередовище пухлини сприяє зв’язування хемокінового рецептора CCR4, який експресується на Treg, до його ліганду CCL22, який секретується багатьма типами пухлинних клітин.[57] Розмноження Treg у місці пухлини також може пояснити зростання їх кількості. Відомо, що цитокін TGF-β, який зазвичай продукується пухлинними клітинами, індукує диференціювання та розмноження T reg клітини. [57]
Білок FOXP3 як фактор транскрипції є важливим молекулярним маркером Treg. Поліморфізм FOXP3 (rs3761548) може брати участь у прогресуванні раку шлунка через вплив на функцію Treg і секрецію імуномодулюючих цитокінів, таких як IL-10, IL-35 і TGF-β.[58]
Загалом, імуносупресія з мікросередовища пухлини значною мірою сприяла невдалим результатам багатьох методів імунотерапії раку. Пригнічення Treg у тваринних моделях призводило до зростання ефективності імунотерапії, і тому багато імунотерапевтичних методів лікування зараз включають пригнічення Treg.[2]
Молекулярна характеристика
Як і інші Т-клітини, Treg розвиваються в тимусі. Останні дослідження показують, що вони визначаються експресією транскрипційного фактора сімейства forkhead FOXP3 (forkhead box protein 3). Експресія FOXP3 необхідна для розвитку Treg і, здається, контролює генетичну програму, яка визначає долю цієї клітини.[59] У переважній більшості Treg, що експресують Foxp3, знаходяться в популяції з головним комплексом гістосумісності (MHC) класу II з обмеженою експресією CD4 (CD4+) і експресують високі рівні альфа-ланцюга рецептора інтерлейкіну-2 (CD25). На додаток до CD4+ CD25+, що експресують FOXP3, існує також незначна популяція Treg, що експресують МНС класу I, CD8+ FOXP3. Ці CD8+ Т-клітини, що експресують FOXP3, не є функціональними у здорових людей, але індукуються при аутоімунних захворюваннях стимуляцією рецепторів Т-клітин для гальмування імунних реакцій, опосередкованих IL-17.[60] На відміну від звичайних Т-клітин, Treg не виробляють IL-2 і тому від початку є анергічними.
У дослідженнях використовується ряд різних методів для виявлення та моніторингу Treg. Спочатку використовували високу експресію поверхневих маркерів CD25 і CD4 (CD4+ CD25+ клітини). Це проблематично, оскільки CD25 також експресується на нерегуляторних Т-клітинах в умовах імунної активації, наприклад, під час імунної відповіді на патоген. Як визначено через експресію CD4 і CD25, Treg становлять близько 5–10% зрілої субпопуляції CD4+ Т-клітин у мишей і людей, тоді як в цільній крові можна знайти приблизно 1–2% Treg. Додаткове вимірювання клітинної експресії білка FOXP3 дозволило провести більш специфічний аналіз Treg (CD4+ CD25+ FOXP3+ клітини). Однак FOXP3 також тимчасово експресується в активованих ефекторних Т-клітинах людини, що ускладнює правильний аналіз Treg з використанням CD4, CD25 і FOXP3 як маркерів у людей. Таким чином, золотим стандартом комбінації поверхневих маркерів Treg в межах неактивованого CD3+ CD4+ є висока експресія CD25 у поєднанні з відсутністю або низьким рівнем експресії поверхневого білка CD127 (IL-7RA). Якщо не цікавить життєздатність клітин, то додавання FOXP3 до комбінації CD25 і CD127 забезпечить додаткову точність дослідження. Було описано кілька додаткових маркерів, наприклад, високі рівні CTLA-4 (цитотоксична молекула, пов’язана з Т-лімфоцитом-4) і GITR (рецептор TNF, індукований глюкокортикоїдами) також експресуються на Treg, однак функціональне значення цієї експресії ще залишається визначити. Існує великий інтерес до виявлення маркерів клітинної поверхні, які унікально та специфічно експресуються на всіх Treg, що експресують FOXP3. Однак на сьогоднішній день така молекула не була ідентифікована.
Ідентифікація Treg після їх активації є складною, оскільки звичайні Т-клітини будуть експресувати CD25, тимчасово експресувати FOXP3 і втрачати експресію CD127 після активації. Було показано, що Treg можна виявити за допомогою маркерного аналізу, індукованого активацією, шляхом експресії CD39[61] у поєднанні з коекспресією CD25 і OX40 (CD134), які визначають антиген-специфічні клітини після 24-48-годинної стимуляції за допомогою антигена. [62]
На додаток до білкових маркерів, в літературі описаний інший метод аналізу та моніторингу Treg. Цей метод заснований на аналізі метилювання ДНК. Тільки в Treg, але не в будь-якому іншому типі клітин, включаючи активовані ефекторні Т-клітини, певна ділянка в межах гена FOXP3 (TSDR, Treg-specific-demethylated region) виявляється деметильованою, це дозволяє виявляти Treg за допомогою реакції ПЛР або інших методів аналізу на основі ДНК.[63] Взаємодія між клітинами Th17 і Treg є важливою при багатьох захворюваннях, таких як респіраторні захворювання.[64]
Останні дані свідчать про те, що тучні клітини можуть бути важливими медіаторами Treg -залежної периферійної толерантності. [65]
Епітопи
Регуляторні епітопи Т-клітин («трегітопи») були відкриті в 2008 році і складаються з лінійних послідовностей амінокислот, що містяться в моноклональних антитілах та імуноглобуліні G (IgG). З моменту їх відкриття, дані вказують на те, що трегітопи можуть мати вирішальне значення для активації природних регуляторних Т-клітин.[66] [67] [68]
Було висунуто гіпотезу про потенційні можливості застосування епітопів Treg: толерантність до трансплантацій і білкових препаратів, гемотрансфузії та діабет I типу, а також зниження імунної відповіді для лікування алергії. [69] [70] [71] [72] [73] [74] [68]
Генетичний дефіцит
Завдяки спадковому захворюванню були виявлені мутації в гені, що кодує FOXP3, були ідентифіковані як у людей, так і у мишей. Це захворювання є найбільш яскравим доказом того, що Treg відіграють важливу роль у підтримці нормальної функції імунної системи. Люди з мутаціями в FOXP3 страждають від важкого аутоімунного розладу з швидким летальним результатом, відомого як синдром імунної дисрегуляції, поліендокринопатії, ентеропатії, пов’язаної з Х-хромосомою (IPEX-синдром).[75]
IPEX-синдром характеризується розвитком системного аутоімунітету на першому році життя, в результаті чого зазвичай спостерігається тріада "водяниста діарея, екзематозний дерматит та ендокринопатія, що найчастіше розглядається як інсулінозалежний цукровий діабет. Більшість людей мають інші аутоімунні явища, включаючи Кумбс-позитивну гемолітичну анемію, аутоімунну тромбоцитопенію, аутоімунну нейтропенію та тубулярну нефропатію. Більшість уражених хлопчиків помирають протягом першого року життя від метаболічних порушень або сепсису. Аналогічне захворювання також спостерігається у мишей зі спонтанною FOXP3-мутацією, відомої як «скурфі» (scurfy).
Посилання
- Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441 (7090): 235–8. May 2006. Bibcode:2006Natur.441..235B. PMID 16648838. doi:10.1038/nature04753. Проігноровано невідомий параметр
|vauthors=(довідка) - Tregs and rethinking cancer immunotherapy. The Journal of Clinical Investigation 117 (5): 1167–74. May 2007. PMC 1857250. PMID 17476346. doi:10.1172/JCI31202. Проігноровано невідомий параметр
|vauthors=(довідка) - Tregs in immunotherapy: opportunities and challenges. Immunotherapy 3 (8): 911–4. August 2011. PMID 21843075. doi:10.2217/imt.11.79. Проігноровано невідомий параметр
|vauthors=(довідка) - Human FoxP3+ regulatory T cells in systemic autoimmune diseases. Autoimmunity Reviews 10 (12): 744–55. October 2011. PMID 21621000. doi:10.1016/j.autrev.2011.05.004. Проігноровано невідомий параметр
|vauthors=(довідка) - Cutting Edge: Regulatory T Cells Facilitate Cutaneous Wound Healing. Journal of Immunology 196 (5): 2010–4. March 2016. PMC 4761457. PMID 26826250. doi:10.4049/jimmunol.1502139. Проігноровано невідомий параметр
|vauthors=(довідка) - Natural and induced T regulatory cells in cancer. Frontiers in Immunology 4: 190. 2013. PMC 3708155. PMID 23874336. doi:10.3389/fimmu.2013.00190. Проігноровано невідомий параметр
|vauthors=(довідка) - Regulatory T cells and treatment of cancer. Current Opinion in Immunology 20 (2): 241–6. April 2008. PMC 3319305. PMID 18508251. doi:10.1016/j.coi.2008.04.008. Проігноровано невідомий параметр
|vauthors=(довідка) - Control of regulatory T cell development by the transcription factor Foxp3. Science 299 (5609): 1057–61. February 2003. Bibcode:2003Sci...299.1057H. PMID 12522256. doi:10.1126/science.1079490. Проігноровано невідомий параметр
|vauthors=(довідка) - Modulation of autoimmune diseases by interleukin (IL)-17 producing regulatory T helper (Th17) cells. The Indian Journal of Medical Research 138 (5): 591–4. November 2013. PMC 3928692. PMID 24434314. Проігноровано невідомий параметр
|vauthors=(довідка) - Regulatory T cells in autoimmmunity*. Annual Review of Immunology 18: 423–49. 2000. PMID 10837065. doi:10.1146/annurev.immunol.18.1.423. Проігноровано невідомий параметр
|vauthors=(довідка) - Naturally occurring regulatory T cells: markers, mechanisms, and manipulation. FASEB Journal 26 (6): 2253–76. June 2012. PMID 22362896. doi:10.1096/fj.11-193672. Проігноровано невідомий параметр
|vauthors=(довідка) - Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annual Review of Immunology 22: 531–62. 2004. PMID 15032588. doi:10.1146/annurev.immunol.21.120601.141122. Проігноровано невідомий параметр
|vauthors=(довідка) - T cell receptor signalling in the control of regulatory T cell differentiation and function. Nature Reviews. Immunology 16 (4): 220–33. April 2016. PMC 4968889. PMID 27026074. doi:10.1038/nri.2016.26. Проігноровано невідомий параметр
|vauthors=(довідка) - Peripheral regulatory T lymphocytes recirculating to the thymus suppress the development of their precursors. Nature Immunology 16 (6): 628–34. June 2015. PMID 25939024. doi:10.1038/ni.3150. Проігноровано невідомий параметр
|vauthors=(довідка) - CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivation-mediated apoptosis of effector CD4+ T cells. Nature Immunology 8 (12): 1353–62. December 2007. PMID 17982458. doi:10.1038/ni1536. Проігноровано невідомий параметр
|vauthors=(довідка) - T-cell tolerance and the multi-functional role of IL-2R signaling in T-regulatory cells. Immunological Reviews 241 (1): 63–76. May 2011. PMC 3101713. PMID 21488890. doi:10.1111/j.1600-065X.2011.01004.x. Проігноровано невідомий параметр
|vauthors=(довідка) - Two subsets of naive T helper cells with distinct T cell receptor excision circle content in human adult peripheral blood. The Journal of Experimental Medicine 195 (6): 789–94. March 2002. PMC 2193736. PMID 11901204. doi:10.1084/jem.20011756. Проігноровано невідомий параметр
|vauthors=(довідка) - Active demethylation of the Foxp3 locus leads to the generation of stable regulatory T cells within the thymus. Journal of Immunology 190 (7): 3180–8. April 2013. PMID 23420886. doi:10.4049/jimmunol.1203473. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Recirculating IL-1R2+ Treg fine-tune intrathymic Treg development under inflammatory conditions. Cellular & Molecular Immunology 18 (1): 182–193. January 2021. PMC 7853075. PMID 31988493. doi:10.1038/s41423-019-0352-8. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|hdl-access=(довідка) - IL-1 receptor 2 (IL-1R2) and its role in immune regulation. Brain, Behavior, and Immunity 32: 1–8. August 2013. PMC 3610842. PMID 23195532. doi:10.1016/j.bbi.2012.11.006. Проігноровано невідомий параметр
|vauthors=(довідка) - Cytotoxic T lymphocyte-associated antigen 4 plays an essential role in the function of CD25(+)CD4(+) regulatory cells that control intestinal inflammation. The Journal of Experimental Medicine 192 (2): 295–302. July 2000. PMC 2193261. PMID 10899916. doi:10.1084/jem.192.2.295. Проігноровано невідомий параметр
|vauthors=(довідка) - The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 450 (7169): 566–9. November 2007. Bibcode:2007Natur.450..566C. PMID 18033300. doi:10.1038/nature06306. Проігноровано невідомий параметр
|vauthors=(довідка) - Interleukin-10 in the regulation of T cell-induced colitis. Journal of Autoimmunity 20 (4): 277–9. June 2003. PMID 12791312. doi:10.1016/s0896-8411(03)00045-3. Проігноровано невідомий параметр
|vauthors=(довідка) - Resolution of airway inflammation and hyperreactivity after in vivo transfer of CD4+CD25+ regulatory T cells is interleukin 10 dependent. The Journal of Experimental Medicine 202 (11): 1539–47. December 2005. PMC 1350743. PMID 16314435. doi:10.1084/jem.20051166. Проігноровано невідомий параметр
|vauthors=(довідка) - Cutting edge: contact-mediated suppression by CD4+CD25+ regulatory cells involves a granzyme B-dependent, perforin-independent mechanism. Journal of Immunology 174 (4): 1783–6. February 2005. PMID 15699103. doi:10.4049/jimmunol.174.4.1783. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - IDO and regulatory T cells: a role for reverse signalling and non-canonical NF-kappaB activation. Nature Reviews. Immunology 7 (10): 817–23. October 2007. PMID 17767193. doi:10.1038/nri2163. Проігноровано невідомий параметр
|vauthors=(довідка) - Expression of ectonucleotidase CD39 by Foxp3+ Treg cells: hydrolysis of extracellular ATP and immune suppression. Blood 110 (4): 1225–32. August 2007. PMID 17449799. doi:10.1182/blood-2006-12-064527. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - T regulatory and primed uncommitted CD4 T cells express CD73, which suppresses effector CD4 T cells by converting 5'-adenosine monophosphate to adenosine. Journal of Immunology 177 (10): 6780–6. November 2006. PMID 17082591. doi:10.4049/jimmunol.177.10.6780. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Role of LAG-3 in regulatory T cells. Immunity 21 (4): 503–13. October 2004. PMID 15485628. doi:10.1016/j.immuni.2004.08.010. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nature Immunology 10 (1): 48–57. January 2009. PMID 19011627. doi:10.1038/ni.1674. Проігноровано невідомий параметр
|vauthors=(довідка) - Cross talk between human regulatory T cells and antigen-presenting cells: Lessons for clinical applications. European Journal of Immunology 51 (1): 27–38. January 2021. PMID 33301176. doi:10.1002/eji.202048746. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Regulatory T cells and immune tolerance. Cell 133 (5): 775–87. May 2008. PMID 18510923. doi:10.1016/j.cell.2008.05.009. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - The emerging role of CTLA4 as a cell-extrinsic regulator of T cell responses. Nature Reviews. Immunology 11 (12): 852–63. November 2011. PMID 22116087. doi:10.1038/nri3108. Проігноровано невідомий параметр
|vauthors=(довідка) - A requisite role for induced regulatory T cells in tolerance based on expanding antigen receptor diversity. Immunity 35 (1): 109–22. July 2011. PMC 3295638. PMID 21723159. doi:10.1016/j.immuni.2011.03.029. Проігноровано невідомий параметр
|vauthors=(довідка) - Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. The Journal of Experimental Medicine 204 (8): 1775–85. August 2007. PMC 2118682. PMID 17620362. doi:10.1084/jem.20070602. Проігноровано невідомий параметр
|vauthors=(довідка) - Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid. Science 317 (5835): 256–60. July 2007. Bibcode:2007Sci...317..256M. PMID 17569825. doi:10.1126/science.1145697. Проігноровано невідомий параметр
|vauthors=(довідка) - Retinoic Acid and Immune Homeostasis: A Balancing Act. Trends in Immunology 38 (3): 168–180. March 2017. PMID 28094101. doi:10.1016/j.it.2016.12.006. Проігноровано невідомий параметр
|vauthors=(довідка) - FOXP3 and the regulation of Treg/Th17 differentiation. Microbes and Infection 11 (5): 594–8. April 2009. PMC 2728495. PMID 19371792. doi:10.1016/j.micinf.2009.04.002. Проігноровано невідомий параметр
|vauthors=(довідка) - Human retinoic acid-regulated CD161+ regulatory T cells support wound repair in intestinal mucosa. Nature Immunology 19 (12): 1403–1414. December 2018. PMC 6474659. PMID 30397350. doi:10.1038/s41590-018-0230-z. Проігноровано невідомий параметр
|vauthors=(довідка) - A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. The Journal of Experimental Medicine 204 (8): 1757–64. August 2007. PMC 2118683. PMID 17620361. doi:10.1084/jem.20070590. Проігноровано невідомий параметр
|vauthors=(довідка) - Induction of stable human FOXP3+ Tregs by a parasite-derived TGF-β mimic. Immunology and Cell Biology 99 (8): 833–847. September 2021. PMID 33929751. doi:10.1111/IMCB.12475. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - A structurally distinct TGF-β mimic from an intestinal helminth parasite potently induces regulatory T cells. Nature Communications 8 (1): 1741. November 2017. PMID 29170498. doi:10.1038/s41467-017-01886-6. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - TGF-β mimic proteins form an extended gene family in the murine parasite Heligmosomoides polygyrus. International Journal for Parasitology 48 (5): 379–385. April 2018. PMID 29510118. doi:10.1016/j.ijpara.2017.12.004. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - The parasite cytokine mimic Hp-TGM potently replicates the regulatory effects of TGF-β on murine CD4+ T cells. Immunology and Cell Biology 99 (8): 848–864. September 2021. PMID 33988885. doi:10.1111/IMCB.12479. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Increase of CD4+CD25highFoxP3+ cells impairs in vitro human microbicidal activity against Mycobacterium tuberculosis during latent and acute pulmonary tuberculosis. PLoS Neglected Tropical Diseases 15 (7): e0009605. July 2021. PMC 8321116. PMID 34324509. doi:10.1371/journal.pntd.0009605. Проігноровано невідомий параметр
|vauthors=(довідка) - Regulatory T Cells As Potential Targets for HIV Cure Research. Frontiers in Immunology 9: 734. 2018. PMC 5908895. PMID 29706961. doi:10.3389/fimmu.2018.00734. Проігноровано невідомий параметр
|vauthors=(довідка) - Nonhuman Primate Testing of the Impact of Different Regulatory T Cell Depletion Strategies on Reactivation and Clearance of Latent Simian Immunodeficiency Virus. Journal of Virology 94 (19): JVI.00533–20, jvi;JVI.00533–20v1. September 2020. PMC 7495362. PMID 32669326. doi:10.1128/JVI.00533-20. Проігноровано невідомий параметр
|vauthors=(довідка) - The therapeutic implications of intratumoral regulatory T cells. Clinical Cancer Research 11 (23): 8226–9. December 2005. PMID 16322278. doi:10.1158/1078-0432.CCR-05-2035. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - CD70+ non-Hodgkin lymphoma B cells induce Foxp3 expression and regulatory function in intratumoral CD4+CD25 T cells. Blood 110 (7): 2537–44. October 2007. PMC 1988926. PMID 17615291. doi:10.1182/blood-2007-03-082578. Проігноровано невідомий параметр
|vauthors=(довідка) - ALS patients' regulatory T lymphocytes are dysfunctional, and correlate with disease progression rate and severity. JCI Insight 2 (5): e89530. March 2017. PMC 5333967. PMID 28289705. doi:10.1172/jci.insight.89530. Проігноровано невідомий параметр
|vauthors=(довідка) - Expanded autologous regulatory T-lymphocyte infusions in ALS: A phase I, first-in-human study. Neurology 5 (4): e465. July 2018. PMC 5961523. PMID 29845093. doi:10.1212/NXI.0000000000000465. Проігноровано невідомий параметр
|vauthors=(довідка) - New Paradigm in the Role of Regulatory T Cells During Pregnancy. Frontiers in Immunology (English) 10: 573. 2019. PMC 6443934. PMID 30972068. doi:10.3389/fimmu.2019.00573. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Decreased maternal serum acetate and impaired fetal thymic and regulatory T cell development in preeclampsia. Nature Communications 10 (1): 3031. July 2019. PMC 6620275. PMID 31292453. doi:10.1038/s41467-019-10703-1. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - The prognostic influence of tumour-infiltrating lymphocytes in cancer: a systematic review with meta-analysis. British Journal of Cancer 105 (1): 93–103. June 2011. PMC 3137407. PMID 21629244. doi:10.1038/bjc.2011.189. Проігноровано невідомий параметр
|vauthors=(довідка) - Suppression, subversion and escape: the role of regulatory T cells in cancer progression. Clinical and Experimental Immunology 171 (1): 36–45. January 2013. PMC 3530093. PMID 23199321. doi:10.1111/j.1365-2249.2012.04657.x. Проігноровано невідомий параметр
|vauthors=(довідка) - Regulatory T Cells in Cancer. Annual Review of Cancer Biology (англ.) 4 (1): 459–477. 9 березня 2020. ISSN 2472-3428. doi:10.1146/annurev-cancerbio-030419-033428. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Cytokine patterns in patients with cancer: a systematic review. The Lancet. Oncology 14 (6): e218–28. May 2013. PMID 23639322. doi:10.1016/s1470-2045(12)70582-x. Проігноровано невідомий параметр
|vauthors=(довідка) - Association of Foxp3 rs3761548 polymorphism with cytokines concentration in gastric adenocarcinoma patients. Cytokine 138: 155351. February 2021. ISSN 1043-4666. PMID 33127257. doi:10.1016/j.cyto.2020.155351. Проігноровано невідомий параметр
|vauthors=(довідка) - Foxp3 occupancy and regulation of key target genes during T-cell stimulation. Nature 445 (7130): 931–5. February 2007. Bibcode:2007Natur.445..931M. PMC 3008159. PMID 17237765. doi:10.1038/nature05478. Проігноровано невідомий параметр
|vauthors=(довідка) - Induced CD8+FoxP3+ Treg cells in rheumatoid arthritis are modulated by p38 phosphorylation and monocytes expressing membrane tumor necrosis factor α and CD86. Arthritis & Rheumatology 66 (10): 2694–705. October 2014. PMID 24980778. doi:10.1002/art.38761. Проігноровано невідомий параметр
|vauthors=(довідка) - Human antigen-specific CD4⁺ CD25⁺ CD134⁺ CD39⁺ T cells are enriched for regulatory T cells and comprise a substantial proportion of recall responses. European Journal of Immunology 44 (6): 1644–61. June 2014. PMID 24752698. doi:10.1002/eji.201344102. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - High levels of human antigen-specific CD4+ T cells in peripheral blood revealed by stimulated coexpression of CD25 and CD134 (OX40). Journal of Immunology 183 (4): 2827–36. August 2009. PMID 19635903. doi:10.4049/jimmunol.0803548. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Quantitative DNA methylation analysis of FOXP3 as a new method for counting regulatory T cells in peripheral blood and solid tissue. Cancer Research 69 (2): 599–608. January 2009. PMID 19147574. doi:10.1158/0008-5472.CAN-08-2361. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Interplay of T Helper 17 Cells with CD4(+)CD25(high) FOXP3(+) Tregs in Regulation of Allergic Asthma in Pediatric Patients. International Journal of Pediatrics 2014: 636238. 2014. PMC 4065696. PMID 24995020. doi:10.1155/2014/636238. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Mast cells are essential intermediaries in regulatory T-cell tolerance. Nature 442 (7106): 997–1002. August 2006. Bibcode:2006Natur.442..997L. PMID 16921386. doi:10.1038/nature05010. Проігноровано невідомий параметр
|vauthors=(довідка) - Tregitope: Immunomodulation Power Tool. EpiVax. 2 серпня 2016.
- Modulation of CD8+ T cell responses to AAV vectors with IgG-derived MHC class II epitopes. Molecular Therapy 21 (9): 1727–37. September 2013. PMC 3776637. PMID 23857231. doi:10.1038/mt.2013.166. Проігноровано невідомий параметр
|vauthors=(довідка) - Activation of natural regulatory T cells by IgG Fc-derived peptide "Tregitopes". Blood 112 (8): 3303–11. October 2008. PMC 2569179. PMID 18660382. doi:10.1182/blood-2008-02-138073. Проігноровано невідомий параметр
|vauthors=(довідка) - New $2.25M infusion of NIH funds for EpiVax' Tregitope, proposed "Paradigm-Shifting" Treatment. Fierce Biotech Research.
- Regulatory T cell epitopes (Tregitopes) in IgG induce tolerance in vivo and lack immunogenicity per se. Journal of Leukocyte Biology 94 (2): 377–83. August 2013. PMC 3714563. PMID 23729499. doi:10.1189/jlb.0912441. Проігноровано невідомий параметр
|vauthors=(довідка) - Application of IgG-derived natural Treg epitopes (IgG Tregitopes) to antigen-specific tolerance induction in a murine model of type 1 diabetes. Journal of Diabetes Research 2013: 621693. 2013. PMC 3655598. PMID 23710469. doi:10.1155/2013/621693. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Teaching tolerance: New approaches to enzyme replacement therapy for Pompe disease. Human Vaccines & Immunotherapeutics 8 (10): 1459–64. October 2012. PMC 3660767. PMID 23095864. doi:10.4161/hv.21405. Проігноровано невідомий параметр
|vauthors=(довідка) - In vitro and in vivo studies of IgG-derived Treg epitopes (Tregitopes): a promising new tool for tolerance induction and treatment of autoimmunity. Journal of Clinical Immunology 33 (1): S43–9. January 2013. PMC 3538121. PMID 22941509. doi:10.1007/s10875-012-9762-4. Проігноровано невідомий параметр
|vauthors=(довідка) - Potential application of tregitopes as immunomodulating agents in multiple sclerosis. Neurology Research International 2011: 256460. 2011. PMC 3175387. PMID 21941651. doi:10.1155/2011/256460. Проігноровано невідомий параметр
|vauthors=(довідка); Проігноровано невідомий параметр|doi-access=(довідка) - Online Mendelian Inheritance in Man IPEX