Хвороби рослин
Хвороби рослин — порушення нормальної життєдіяльності рослинного організму, яке проявляється в розладі його фізіологічних функцій і структури. Хвороби рослин призводять до ураження окремих органів або загибелі всієї рослини, отже — до зниження урожаю і якості рослинницької продукції. Причиною Хвороби рослин можуть бути патогенні чинники (віруси, бактерії та інші організми) й несприятливі умови середовища. Розрізняють інфекційні і неінфекційні хвороби рослин. Інфекційні хвороби рослин залежно від збудників, поділяють на грибні хвороби рослин, бактеріальні хвороби рослин, вірусні хвороби рослин, нематодні хвороби рослин і хвороби, збудниками яких є рослини-паразити (Омела, Вовчок, Повитиця тощо).
| Сільське господарство |
|---|
 Борошниста роса на листі гарбуза |
|
|
|
Технологія
|
|
|
Причини
Неінфекційні хвороби рослин виникають внаслідок високої або низької температури, різких коливань температури повітря й ґрунту (наприклад, вигоряння посівів, вимерзання рослин, випирання рослин); внаслідок надлишку або нестачі вологи (зокрема, вимокання посівів, стікання рослин, передчасне в'янення рослин); порушення режиму мінерального живлення рослин, неправильного співвідношення елементів живлення в ґрунті та інше. Залежно від локалізації розрізняють місцеві і загальні хвороби рослин. Місцеві хвороби рослин характеризуються ураженням окремих органів рослини у вигляді плямистостей (церкоспороз буряків, картоплі, септоріоз злаків, коколікоз вишні і черешні), пустул (іржа рослин), порошкоподібного або павутинного нальоту (Борошниста роса, Несправжня борошниста роса), наростів (пухирчаста сажка кукурудзи). При загальних хворобах рослин, найчастіше спостерігається в'янення рослин в цілому внаслідок ураження судинної системи (наприклад, фузаріози льону, вілт бавовника, картоплі, соняшника тощо). За характером прояву, виділяють латентні (приховані) хвороби рослин, які протікають без симптомів (деякі вірусні, мікоплазмові і бактеріальні хвороби); хронічні хвороби рослин, коли рослина хворіє тривалий час або увесь період життя (гниль деревини, чорний рак яблуні). Хвороби рослин передаються з насінням і садивним матеріалом, рослинними рештками, через ґрунт, інвентар, комахами, вітром, водою тощо.
У непорушених людиною фітценозах шкода трав'янистим та нетрав'янистим рослинам, яку наносять гриби, є невеликою, більш того, вони виконують важливі функції регуляторів чисельності компонентів фітоценозів. Інша картина спостерігається у посівних сільскогосподарських культур (агроценозах) й лісних масивах, які зазнали антропогенного впливу. Такі рослинні спільноти часто зазнають масових захворювань (епіфітотій), які, якщо не проводити спеціальних захисних заходів (посів й посадка стійких сортів, обробка насіння й вегетуючих рослин хемічними речовинами - фунгіцидами й ін.), можуть викликати гибель усієї популяції.
Причини масових хворіб пов'язані із порушенням людиною природних чинників, регулюючих взаємовідносини рослин й їх паразитів.
- Замість складних багатовидових фітоценозів, у яких випадково розподілені по ступені сприйняття до того чи іншого гриба види, у сільськогосподарських посівах й лісових посадках переважають чисті культури одного виду або навіть сорти рослин, на яких вірулентний для них вид або штам паразиту розвивається із швидкістю лісної пожежі.
- Разом із рослинами, інтродукованими у нові регіони, туди попадають й їхні паразити, які можуть перейти на місцеві рослини, які не мають вироблених у процесі коеволюції механізмів стійкості до цих паразитів.
- Орна культура, застосування добрив й інші агротехнічні прийоми привели до корінних змін умов існування ґрунтових мікроорганізмів. Впала чисельність безидіальних грибів, міцелій яких інгібує розвиток сумчастих й недосконалих грибів - основних паразитів рослин. Останні не лише збільшили свою чисельність, але й підвищили патогенні властивості по відношенню до рослин, стали збудниками гниття коріння й інших хворіб рослин.
- Високе рекреаційне навантаження на лісові масиви (витоптування й ущільнення ґрунту, що призводить до зниження аерації коріння), наявність шкідливих речовин (ксенобіотиків) у повітрі й ґрунті знижують природні механізми стійкості до хворіб.
Масові хвороби рослин призводили до голоду й гибелі населення у місцях, де вирощувалася лише одна харчова культура. Наприклад, у 40-х роках 19 ст. від неурожаю картоплі, викликаного фітофторозом, померло й імігрувало за океан більшість населення Ірландії, а через сто років подібна біда (гибель 2 мільйонів жителів) сталася у Бенгалії, внаслідок гибелі рису від гельмінтоспоріозного опіку листя. Грибні хвороби рослин, зокрема іржавіння, зіграли вирішальну роль у переміщенні центру вирощування кави з Південно-Східної Азії (Індії, Цейлону) до Південної Америки. На західному узбережжі США рак кашанів, викликаний сумчастим грибом Cryphonectria parasitica, призвів до зміни ландшафтів: каштанові ліси замінилися кущовими зарослями[1].
Самозахист рослин (хімічний поєдинок)
У процесі еволюції у рослин виробилися захисні функції. Наприклад, часник має у складі речовини із фітонцидними властивостями (наприклад, хлорелін, томатин, сатівін). Аспектом міжвидової взаємодії рослин та мікроорганізмів є стресові метаболіти рослин, або фітоалексини. Фітоалексини - антибіотичні речовини, які утворюються у тканинах рослин, коли до них попадає мікробна інфекція. Їхній біосинтез індукується або різко інтенсифікується лише при контакті клітин рослини-господаря із мікроорганізмом, тобто в результаті взаємодії двох метаболічних систем. Захисна реакція спостерігається лише у живих тканин, а їх розвиток призводить до некрозу, тобто фітоалексини - це продукти некробіозу клітини господаря. Антибіотична дія фітоалексинів обумовлена тим, що у високій концентрації вони порушують центральні процеси обміну - біосинтез нуклеїнових кислот та білка, дихання, окисне фосфорилювання, та змінюють проникність клітинних мембран; хоча фітоалексини не специфічні по відношенню до грибів, останні можуть бути чутливими до них у різній степені. Стійкість рослини до дії патогенів визначається швидкістю накопичення фітоалексинів в ураженій ділянці: чим швидше вони там накопичуються, тим стійкішою є рослина по відношенню до даного мікроорганізму, швидкість цієї реакції є генетично детермінованою. Утворення фітоалексинів обмежується лише ураженою тканиною й тканинами, які оточують осередок інфекції. Сутність процесу - відгородження частини заради спасіння цілого.
Фітоалексини рослин, які відносяться до різних класів є несхожими між собою, але всередині сімейства вони зазвичай належать до близьких структурних типів. Для бобових (гороху, фасолі, вики, клеверу, люцерни) характерні геероциклічні сполуки - похідні птерокарпану: пізатин, гомопізатин, медикарпін, глюкозид трифоліризин, фасеолін тощо. У Vicia fava, крім того, у відповідьна зараження Botrytis cinerea або Botrytis Fabae виробляються ацетиленові фітоалексини, наприклад вієронепоксид. У коріннях моркви у відповідь на мікробну інфекцію виробляються 6-метоксимеллеїн, У бульбі орхідеї - орхінол, у бульбі атату - іпомеамарон. Із стебла зараженої вілтом бавовни виділений ізогемігосипол, який є фітоалексином, комбінуючим у собі фенольну та секвітерпенову природу. Для пасльонових у відповідь на мікробну інфекцію в уражених тканинах відбувається перебудова метаболізму ізопреноїдів: синтез звичайних для здорової рослини тритерпенів й стероїдних алкалоїдів поступається інтенсивному біосинтезу сесквітерпенових фітоалексинів. З картоплі, ураженої фікоміцетом Phytophtora infestans, виділені лубімін, оксілубімін, солаветівон, фітуберин й ризитин; усі вони мають сильну антифунгальну дію. Картопля та інші рослини сімейства пасльонових (Salanacteae) піддається фітофторозу - захворюванню, яке викликається грибами Phytophtora. Деякі сорти картоплі є стійкими до інфекції, оскільки відповідають виробленням фітоалексинів, наприклад, з сорту "Ріширі" був виділений фунгітоксичний метаболіт, який назвали рішитином. У табака у інфікованих листях з'являється квієзон - потужний інгібітор проростання конідій у Perenospora tabacina. У дурману Datura stramonium окрім лубіміну й оксилубіміну утворюється гермакрендіол. Для винограду й інших представників Viticaceae фітоалексинами є резвератрол й його циклоолігомери: димер ε-вініферин, тример α-вініферин тощо. Соя (Glycine max L.Merrill) виробляє фітоалексин, який накопичується у місцях травм тканин або при інфекції. При уражені сосни виробляється піостробін. У листях евкаліпту був знайдений робустаол як компонент протималярійного комплексу речовин. Хемічну природу діфенілметанів мають також деякі антибактеріальні інгридієнти звіробоїв (Hypericum). Наприклад, друмондіни з H.drummondii по активності проти грампозитивних бактерій переважають антибіотик стрептоміцин. У лусці цибулі міститься пірокатехін, що дозволяє протистояти шкідникам. В цибулі міститься антибіотик алін[2]. Бензойна кислота є фітоалексином яблуні, який виробляється у її листях у відповідь на інфікування дерева грибком Nectria galligena. [3] Якщо придивитися до листя томату, то можна побачити, що вони покриті трихомами. Коли на листя сідає комаха або коли вона намагається поїдати лист, трихоми руйнуються й вивільняється 2-тридеканон, який є токсичним для комах (такі метаболіти називають детеррентами або антифідантами). У листях томату міститься глікоалкалоїд томатин[4].
Рослина крокіс (Cortamus tinctorius) відповідає на зараження патогенними грибами виробленням триацетиленового фітоалексину сафінолу. Водорості дінофлагеллати Gymnodinium breve виробляють сильні бреветоксини, що є бідством для прибережних районів, коли масово гине риба й отруюються люди (вода при цьому набуває червоного забарвлення у прибережних районах - подібно до того, як Ceratium hirundinella надає буроватий відтінок воді в Україні). Дінофлагеллат виробляє палітоксин й маїтотоксин (маїтотоксин є найбільшою з відомих природних мономерних молекул, він у 200 000 разів більш отруйний, ніж цианіди натрію та калію).
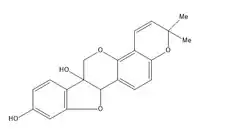
Стійкість дерева до гниття визначається наявністю у ньому стильбенів із антибактеріальними й протигрибковими властивостями. Наприклад, деревина сосни містить піносильвін, а евкаліпти багаті резвератролом. Резвератрол у листях арахісу виконує функцію фітоалексину.
У ході хемічного поєдинку із рослиною-господарем патогенні гриби здатні перетворювати фітоалексини у менш токсичні для них продукти. Наприклад, птерокарпанові алексини з солодкого й червоного клеверу метаболізуються трьома патогенними грибами - Botrytis cinerea, Colletotrichum cofeanum та Colletotrichum lindemuthianum - у менш токсичні сполуки за рахунок гіродксилювання й метилювання-деметилювання, а вієронепоксид відновлюється під дією B.cinerea у вієролепоксид, який має меншу антифунгальну активність. [5]
Запобігання
Заходи боротьби спрямовані насамперед на запобігання хворобам: дотримання законів землеробства і агротехніки, створенням цим самим найкращих умов для росту і розвитку сільськогосподарських культур, вирощування стійких до хвороб сортів і гібридів, створення здорового посівного і садивного матеріалу, застосування біологічних, фізичних, механічних і хімічних засобів захисту рослин, додержання карантинних заходів.
При промисловому виробництві посадочного матеріалу головною проблемою є низкий процент виживання на етапі кліматичної інтеграції й слабкий ріст адаптованих мікророслин після висадження у відкритий ґрунт. Покращити мінеральне живлення й підвищити стійкість рослин до водного стресу може використання симбіотичних мікроорганізмів, які присутні у природних умовах проростання.
Мікоризація
Мікоризація є зараженням рослин культурами мікоризних грибів. Цей прийом підвищує приживання, урожайність та стійкість рослин. Більшість видів наземних рослин (80-90%) здатне до симбіотичної взаємодії із грибами відділу Glomeromycota, з представниками якого рослини утворюють арбускулярну мікоризу (АМ). Утворюючи симбіоз із корінням рослин, гіфи гриба збільшують площу абсорбції кореневої системи, завдяки чому у декілька разів підвищується всмоктувальна здатність, а мінеральні речовини надходять всередину кореня у легкозасвоюваній формі. Мікоризація коріння рослин може пригнічувати ріст й розвиток корневих патогенних мікроорганізмів: ґрунтових грибів й бактерій. Після інокуляції АМ підвищувалася стійкість томату до зараження Fusarium oxysporum та Pseudomonas syringae, a казуарини - до зараження Fusarium vesicibesum. Розрізняють специфічні й неспецифічні механізми пригнічення мікоризними грибами патогенних організмів. Неспецифічні ефекти пов'язані із покращенням мінерального живлення рослини-господаря. Специфічна дія заснована на змінах, які зачіпають склад ризосферної мікрофлори, секреторну активність й анатомію (лігніфікація й суберинізація тканин) коріння, реалізацію захисних реакцій рослини[6].
АМ-гриби розповсюджені усюди, вони добре розвиваються у широкому діапазоні кислотності ґрунтів, температури, вологості та аерації. Арбускулярна мікориза здійснює загальностимулюючу дію на рослини, збільшує активність фотосинтезу, покращує водний статус й мінеральне живлення, впливає на фітогормональний статус, підвищує стійкість рослин до патогенів. АМ-гриби сприяють переведенню живильних речовин, зокрема фосфору та мікроелементів, у форми, доступні для рослин.
| Види АМ-рибів | Рослини |
|---|---|
| Glomus epigaeum | Яблуня |
| Glomus fasciculatum, G. caledonium, G. monosporum,
G. constrictum, G. occultum, G. vesiforme, G. mosseae, Gigaspora margarita |
Ківі (Actinidia deliciosa),
Виноградна Лоза (Vitis sp.) |
| Glomus aggregatum, G.deserticola | Черешня (Prunus avium),
Спірея (Spiraea vulgaris), Бузок (Syringa japonica) |
| Glomus fasciculatum, G. intraradices | Фініковая пальма (Phoenix dactylifera) |
| Glomus mosseae, G. coronatum, G. caledonium | Алича (Prunus cerasifera) |
| Glomus intraradices, G. geosporum | Суниці (Fragaria ananassa) |
| Glomus intraradices, G. macrocarpum, G. mosseae | Суниці (Fragaria ananassa, F. virginiana, F. vesca) |
| Glomus intraradices, G. fasciculatum, G. caledonium, G. monosporum | Черешня (Prunus avium) |
Бактеризація
Метод бактеризації є не менш ефективним при клональному розмноженні, ніж мікоризація. Як і за мікоризації, симбіотичні бактерії розвиваються на живильному середовищі лише у присутності рослини й не потребують повторної інокуляції. Бактерії роду Pseudomonas стимулюють рыст рослин in vitro, покращують водний обмін, мінеральне живлення, а також підвищують стійкість до патогенних організмів при пересаджуванні. Для бактеризації частіше використовують стимулюючі ріст бактерії родів Pseudomonas, Bacillus, Azospirillum, Azotobacter та інші. Зараження рослин можна проводити як in vitro, так й ex vitro[7].
Див. також
Посилання
- Довідник хвороб //Аграрна енциклопедія
- Белякова Г.А.и др. - Ботаника, т.1.
- Беккер М.Е. - Введение в биотехнологию.
- А.А.Семенов - Очерк химии природных соединений.
- З.Гауптман, Ю.Грефе, Х.Ремане - Органическая химия.
- М.Барбье - Введение в химическую экологию.
- Николай Битюцкий - Минеральное питание растений.
- М.И. Яблонская, М.С. Гинс, М.А. Молчанова - БИОТИЗАЦИЯ РАСТЕНИЙ IN VITRO.