Альбатросові
Альбатро́си, або альбатро́сові (Diomedeidae) — родина морських птахів ряду буревісникоподібних (Procellariiformes), широко розповсюджених у Південному океані (води навколо Антарктиди) і в північній частині Тихого океану. На півночі Атлантики вони відсутні, проте на підставі викопних залишків можна припустити, що раніше вони зустрічалися і там. Альбатроси є одними з найкрупніших літаючих птахів, а у мандрівного (Diomedea exulans) і південного королівського альбатросів (Diomedea epomophora) розмах крил досягає рекордної величини — до 3,7 м і навіть більше[1]. У забарвленні оперення у більшості альбатросів переважають чорні, білі й бурі кольори. Ніздрі містяться по боках дзьоба, кожна в окремій трубочці. Крила вузькі й довгі. Молоді птахи набирають вигляду дорослих особин на 4—5-й рік життя. Зазвичай всіх альбатросів поділяють на 4 роди, але з приводу кількості видів серед орнітологів є певні розбіжності.
? Альбатросові | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Північний королівський альбатрос (Diomedea sanfordi) | ||||||||||||
| Біологічна класифікація | ||||||||||||
| ||||||||||||
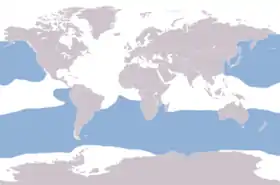 Ареал альбатросових (блакитний) | ||||||||||||
Роди | ||||||||||||
| ||||||||||||
Посилання | ||||||||||||
| ||||||||||||
Альбатросові комфортно відчувають себе в повітрі, уміло використовуючи динамічне і похиле ширяння для подолання значних відстаней з малою енергією. Живляться кальмарами, рибою, крилем, полюючи на поверхні моря або пірнаючи за здобиччю з повітря. Розмножуються в колоніях, в основному влаштовуючи свої гнізда на віддалених океанічних островах, часто спільно з іншими видами птахів. Моногамні, формування пари між самцем і самкою займає досить багато часу — зазвичай декілька років, при цьому залицяння супроводжується шлюбними танцями. Після утворення пара зберігається протягом всього життя. Сезон розмноження і виховання може тривати більше року, починаючи з кладки яєць і закінчуючи першим польотом молодих пташенят. У кладці буває тільки одне яйце.
З 21 виду альбатросових, що визнаються Міжнародним союзом охорони природи, 19 знаходяться під захистом цієї організації як види, яким більшою чи меншою мірою загрожує небезпека вимирания. У минулі століття популяція цих птахів різко знизилася унаслідок їх масового винищування заради пір'я для дамських капелюшків, а у наш час їх існуванню загрожують інтродуковані (тобто невластиві даній місцевості, завезені) тварини, такі як здичавілі коти, які полюють за яйцями, пташенятами і гніздовими птахами; забруднення навколишнього середовища; виснаження рибних запасів моря і загибель у риболовецьких снастях при ярусному лові. Високою є смертність птахів при рибному лові на гачок з наживкою, яка приваблює птахів. Урядові і природоохоронні організації розробляють заходи для зменшення цієї загрози.
Еволюція
У своїх роботах Сіблі і Алквіст на підставі вивчення адаптивного розходження пришли до висновку, що ряд буревісникоподібних, до якого входять альбатроси, бере свій початок в олігоцені (35—20 млн років тому). Проте, можливо ця група з'явилася ще раніше: скам'янілі рештки птахів роду Tytthostonyx, який зазвичай асоціюється з буревісникоподібними, відносяться до крейдяного періоду (70 млн років тому). Молекулярний аналіз показує, що першими від єдиного стародавнього стовбура відійшли буревісники (Procellariidae), потім альбатроси, а пізніше — качуркові (Hydrobatidae) і пірнаючі буревісники (Pelecanoididae). Найбільш ранні викопні знахідки альбатросів належать до еоцену, хоча деякі з них віднесені до родини лише ймовірно — з сучасними птахами вони мають мало спільного. Серед знахідок можна назвати роди Murunkus (середній еоцен, Узбекистан), Manu (ранній олігоцен, Нова Зеландія) і одну неописану форму (пізній олігоцен, Південна Кароліна, США). Крім того, ще одна скам'янілість — Plotornis (середній міоцен, Франція), раніше віднесена до буревісників, у наш час[коли?] також розглядається як представник альбатросів. В останньому випадку знахідка відноситься до середнього міоцену, коли сучасні роди альбатросів вже сформувалися. Про це свідчать рештки видів Phoebastria californica і Diomedea milleri, що відносяться до середнього міоцену (15 млн років тому) і виявлені в Каліфорнії, США. Аналогічні знахідки в південній півкулі доводять розбіжність родів Phoebetria і Thalassarche близько 10 млн років тому[2].
У північній півкулі відомо більше викопних знахідок альбатросів, ніж у південній. Багато форм виявлено також і в північній частині Атлантичного океану, де в наш час[коли?] альбатроси відсутні, причому більшість знахідок відносять до роду Phoebastria. Наприклад, залишки стародавньої колонії білоспинного альбатроса (Phoebastria albatrus) були відкриті на одному з Бермудських островів[3]. Крім того, залишки виду Phoebastria anglica були виявлені у відкладеннях Північної Кароліни (США) і Англії.
Загальна характеристика
Опис

Альбатроси — великі птахи, найбільші серед всіх буревісникоподібних. Дзьоб — великий, могутній і з гострими краями; наддзоб'я на кінці гачкувато заломлено. Дзьоб складається з декількох рогових пластинок і має пару довгих трубочок-ніздрів, що дали одну з назв ряду (трубконосі). На відміну від інших буревесникоподібних, у альбатросів ніздрі розташовані з боків дзьоба, а не у верхній його частині. Завдяки таким ніздрям у альбатросів є гострий нюх, що рідко зустрічається у птахів, і, як і представники інших споріднених родин, альбатроси шукають джерела корму за запахом[4]. На ногах задній палець відсутній, а три передні пальці зв'язано суцільними перетинками. Лапи сильні в порівнянні з іншими трубконосими: зі всього ряду тільки альбатроси і південний гігантський буревісник (Macronectes giganteus) здатні добре пересуватися суходолом.
Оперення дорослих птахів у більшості видів зазвичай має різні варіації чорного верху і зовнішньої частини крил і білих грудей. Забарвлення птахів варіює від майже повністю білого, як у самців південного королівського альбатроса (Diomedea epomophora), у яких темними залишаються лише кінчики і краї крил, до темно-бурого, як в острівного альбатроса (Diomedea amsterdamensis), пір'я якого, в основному, темно-буре, а на грудях є виразна темно-коричнева смуга. У деяких видів з родів Thalassarche і Phoebastria на голові є особливі відмітини на зразок плям навколо очей чи сірі або жовті плями на потилиці. Три види альбатросів: (чорноногий альбатрос (Phoebastria nigripes), темноспинний димчастий альбатрос (Phoebetria fusca) і світлоспинний димчастий альбатрос (Phoebetria palpebrata)) — значно відрізняються від інших птахів, їх оперення майже повністю темно-коричневе або темно-сіре (у світлоспинного димчастого альбатроса). У молодих птахів оперення декілька відрізняється від оперення дорослих особин і їм потрібно кілька років для придбання зрілого забарвлення.
Розмах крил у деяких видів з роду Diomedea, зокрема, у мандрівного і південного королівського альбатросів, найбільший серед всіх видів птахів, що живуть нині, і може перевищувати 340 см. У решти видів розмах крил помітно менший. Крила жорсткі, довгі, вузькі і дугоподібні, з потовщеною передньою кромкою обтічної форми. Птахи здатні долати значні відстані, і при польоті використовують один з двох можливих технічних прийомів: динамічне або похиле ширяння. При динамічному ширянні вони отримують енергію вертикального градієнту вітру, переміщаючись у горизонтальних повітряних потоках з різною швидкістю вітру, і тим самим економлять власні сили. При похилому ширянні альбатроси набирають висоту при зустрічному вітрі і потім ширяють у зворотному напрямі. Відносна дальність ширяння альбатросів становить від 1:22 до 1:23, тобто, при опусканні на кожен метр висоти вони в змозі пролетіти 22—23 метри. При наборі висоти використовують плечові сухожилля, які при розтяганні фіксують крило і дозволяють утримувати його без м'язевих зусиль — (адаптація властива альбатросам і гігантським буревісникам)[5].

Окрім застосування певної техніки ширяння, альбатроси використовують і передбачені метеоролігічні умови: у південній півкулі при русі на північ убік від гніздової колонії вони відхиляються за годинниковою стрілкою, а у зворотний бік — проти неї[6]. Вони так добре адаптувалися до повітряного способу життя, що серцевий ритм у польоті близький до фонової частоти ударів серця у стані спокою. Така ефективність досягається за рахунок того, що основні витрати енергії під час пошуку їжі витрачаються не на подолання відстані, а на зліт, посадку і полювання[7]. Маючи чудові здібності до тривалого польоту, альбатроси долають величезні відстані у пошуках широкої розкиданої кормової бази і при цьому витрачають невелику кількість енергії. Будова крил дозволяє знаходитися в повітрі тривалий час, але робить альбатросів залежними від вітру і хвиль і недостатньо пристосованими до активного польоту. У більшості видів мускулатура розвинена недостатньо для тривалого махового польоту. У безвітрі птахи вимушені тривалий час відпочивати на поверхні моря до тих пір, поки порив вітру не допоможе їм злетіти. Сплять вони також на морі, а не в повітрі, як це колись вважалося. У видів роду Phoebastria є ще один спосіб польоту: вони ривком роблять змах крилами, а потім плавно ширяють у повітрі[8].
Розповсюдження
Більшість альбатросів мешкають у південній півкулі від Антарктиди на півдні до Австралії, Південної Африки і Південної Америки на півночі. Виключенням є чотири види з роду Phoebastria, три з яких поширені в північній частині Тихого океану від Гавайських островів до Японських, Каліфорнії і Аляски, а один — галапагоський альбатрос (Phoebastria irrorata) — гніздиться на Галапагоських островах і годується у тихоокеанського узбережжя Південної Америки. Розповсюдження переважно на високих широтах пояснюється морфологічними особливостями, які вимагають наявність вітру при ширянні. Слабка здібність до активного махового польоту робить важкоздійсненним завдання перетину екваторіальної штильової смуги. Винятком є галапагоський альбатрос, який користується вітрами, що викликаються холодною океанічною течією Гумбольдта.

Причини, за якими альбатроси вимерли в північній Атлантиці, точно невідомі. Проте вважають, що колонія білоспинних альбатросів на Бермудських островах зникла унаслідок теплого міжльодовикового періоду, що викликав затоплення місць гніздування[3]. Представники деяких південних видів іноді випадково залітають на північ і залишаються там протягом десятиліть. Один з таких «вигнанців», альбатрос Thalassarche melanophris, протягом багатьох років повертався в колонію північних олуш (Morus bassanus) в Шотландії в надії знайти там собі пару[9].
Використання супутникового спостереження дає ученим-орнітологам хорошу картину пересування альбатросів в океані. Вони не здійснюють сезонних міграцій, проте після закінчення періоду розмноження розосереджуються у великому діапазоні та — у разі південних видів — часто здійснюють приполярні навколосвітні подорожі[10]. Є свідоцтво того, що у різних видів все ж таки є розділення природних зон. Порівняння екологічних ніш двох близьких видів — кемпебеллівського (Thalassarche impavida) і сіроголового альбатросів (Thalassarche chrysostoma), що гніздяться на островах Кемпбелл у Новій Зеландії, показало, що в першому випадку птахи годуються в районі мілин острова Кемпбелл, а в другому — в пелагічній океанічній зоні. Мандрівний альбатрос також шукає собі прожиток строго у водах з глибиною більше 1000 м, що дало привід одному ученому відмітити: «Таке враження, що птахи бачать знак „Входу немає“ там, де глибина зменшується до менше 1000 м» (англ. «It almost appears as if the birds notice and obey а 'No Entry' sign where the water shallows to less than 1000 m»)[2]. Крім того, різні зони проживання спостерігаються навіть у самок і самців одного виду: дослідження у відношенні тристанського альбатроса (Diomedea dabbenena) показало, що, розмножуючись на острові Гоф (англ. Gough Island), самці після шлюбного періоду відлітають на захід від нього, а самки — на схід[2].
Живлення

Основу раціону альбатросів складають головоногі молюски, риба і ракоподібні, хоча при нагоді вони живляться зоопланктоном або падлом[6]. Варто відзначити, що для більшості видів вичерпна інформація зібрана лише під час розмноження, коли альбатроси повертаються з полювання до місць гніздування, і може не враховувати інші періоди. Значення кожного з перерахованих джерел їжі може варіювати у різних видів, і навіть у різних популяцій одного виду. Деякі птахи в основному полюють на кальмарів, інші надають перевагу крилю або рибі. З двох видів, що мешкають на Гавайських островах, один — чорноногий альбатрос (Phoebastria nigripes) в основному живляться рибою, а другий — темноспинний альбатрос (Phoebastria immutabilis) — переважно кальмарами.
На підставі показників вбудованих датчиків, які записують час заковтування води і тим самим дають знати час прийому їжі, було висловлено припущення, що альбатроси шукають собі корм переважно в денний час доби. Аналіз залишків кальмарів, яких відригнули птахи, показав, що багато кальмарів були занадто великими, щоб їх можна було зловити живими[11]. Крім того, в шлунках були знайдені залишки організмів теплих широт, куди альбатроси навряд чи могли залетіти. На підставі отриманих даних був зроблений висновок, що у деяких видів, наприклад, мандрівного альбатроса, падло може займати значну частину раціону. Джерело знайдених мертвих кальмарів є предметом дискусій: деякі вважають, що вони можуть бути покидьками рибальства, що померли під час нересту, або були викинуті іншими організмами, що живляться цими тваринами — переважно китоподібними — наприклад, кашалотами (Physeter macrocephalus), короткоплавцевими гриндами (Globicephala macrorhynchus) і високочолими пляшконосами (Hyperoodon ampullatus). Інші види, такі як чорнобривий або сіроголовий альбатроси, живляться дрібнішими видами кальмарів, які у разі загибелі зазвичай тонуть. Отже, для цих птахів падло не грає таку значну роль в їх раціоні.
До недавнього часу вважалося, що альбатроси знаходять собі прожиток переважно на поверхні моря, плаваючи і хватаючи кальмарів, рибу та інший корм, викинутий на поверхню океанічною течією, хижаками або внаслідок загибелі. Як експеримент птахам упровадили капілярні пристрої, що реєструють максимальну глибину, на яку вони занурюються при пірнанні. Результати дослідження показали, що деякі види, наприклад, мандрівні альбатроси, не упірнають більш ніж на метр; а інші, як димчасті альбатроси, зазвичай занурюються на глибину до 5 м, проте здатні упірнути і ще глибше, до 12,5 м[12]. Крім того, птахи можуть упірнути за жертвою не тільки з поверхні моря, але і з повітря[13].
Розмноження

Альбатроси розмножуються в колоніях, зазвичай на ізольованих островах. Якщо острів великий, то вони віддають перевагу крутим і скелястим мисам з хорошим доступом до моря з різних напрямків. Прикладом такої колонії може служити півострів Отаго в районі міста Данідін у Новій Зеландії. Колонії можуть бути як дуже великими і густонаселеними (наприклад, щільність гніздів'їв чорнобривого альбатроса на Фолклендських островах досягає 70 гнізд на кожні 100 м²), так і дрібнішими і сильно розрідженими, як це властиво альбатросам родів Diomedea і Phoebetria. Історично на всіх островах, на яких гніздяться птахи, наземні ссавці відсутні. Альбатроси дуже філопатрічні, тобто, мають тенденцію повертатися на те саме місце, де вилупилися самі. Наприклад, за результатами дослідження темноспинного альбатроса (Phoebastria immutabilis) був зроблений висновок, що відстань між гніздом пташеняти і його першим власним гніздом у середньому становить 22 м[14].
Альбатросам, як і більшості інших морських птахів, притаманна типова K-стратегія, тобто вони живуть набагато довше за інших птахів, розмножуються в зрілішому віці і значний час присвячують вихованню потомства. Максимальна тривалість життя альбатросів у природі може досягати 50 років; найстаріший відомий північний королівський альбатрос (Diomedea sanfordi) був окільцьований у зрілому віці й далі спостерігався протягом 51 року, що дало привід ученим визначити його приблизний максимальний вік у 61 рік[15].
Статева зрілість у птахів настає досить пізно, приблизно через 5 років життя. Проте, вже досягнувши статевої зрілості, альбатроси не починають розмножуватися ще протягом ряду років (до 10 років у деяких видів). Молоді птахи відвідують колонію перш, ніж приступити до розмноження, і витрачають декілька років на залицяння за майбутнім партнером, при цьому ці залицяння супроводжуються шлюбними ритуалами і танцями[16]. Повертаючись у колонію вперше, птахи вже показують стереотипну поведінку, яка і формулює характерну пташину мову, проте ще не в змозі ні визначити таку ж поведінку у інших птахів, ні відповісти на неї адекватно[6]. Пройшовши шлях проб і помилок, молоді птахи починають розуміти її «синтаксис» і досягають досконалості в процесі залицяння і шлюбних ритуалів. Сприйняття чужої поведінки молодняком відбувається швидше в оточенні зрілих птахи.

Шлюбні танці є координованим виконанням різних дій, які можуть включати чищення дзьобом власного пір'я, поглядання по сторонах, спів, клацання дзьобом, наїжачування і будь-яку комбінацію вище перерахованого[17]. Повернувшись до колонії вперше, птахи прагнуть залицятися до багатьох потенційних партнерів, але протягом кількох років їх кількість незмінно падає до тих пір, поки не залишається тільки один партнер, і пара нарешті формується. Самець і самка продовжують спілкуватися між собою своєю мовою, яка, кінець кінцем, стає властивою виключно цій парі. Сформувавши пару на все життя, птахи вже ніколи не повертаються знов до ритуальних танців. Вважають, що ретельно і скрупульозно виконуючи шлюбні ритуали, птахи переконуються в правильності вибору партнера, оскільки сумісне висиджування яєць і доглядання за пташенятами вимагає великих фізичних витрат і тривалого часу. Навіть ті види, в яких цикл відтворення займає менше року, дуже рідко знову відкладають яйця наступного року[2]. У птахів роду Diomedea, наприклад, у мандрівного альбатроса, період між кладкою яєць і першим польотом пташеняти займає більше року. Альбатроси відкладають тільки одне яйце за сезон; якщо з якої-небудь причини кладка не відбулася (знищена хижаком, ненавмисно розбилася), то самка здатна в той же рік відкласти повторно. Пари розпадаються рідко — зазвичай це може відбутися після декількох неуспішних спроб обзавестися потомством.
Самці першими прилітають на місце гніздування, де і чекають своїх самок. Альбатроси південної півкулі будують великі гнізда, тоді як у північних видів вони примітивніші. Галапагоський альбатрос (Phoebastria irrorata) гнізд не будує зовсім і навіть катає своє яйце територією колонії на відстань до 50 м, що іноді приводить до його втрати[18]. Як матеріал для гнізда використовуються купки рослин або насип із землі або торфу. У всіх видів як самець, так і самка беруть участь у насиджуванні яєць, безперервна вахта одного з батьків може займати від одного дня до трьох тижнів. Інкубаційний період становить 70—80 днів (чим більший птах, тим довший період), що є рекордом за тривалістю серед всіх птахів. У період насиджування птахи втрачають до 83 г своєї ваги за день[19].

Пташенят, що вилупилися, ще протягом біля трьох тижнів висиджують і охороняють до тих пір, поки вони не стають досить великими, щоб зуміти зігрітися самостійно і у разі потреби постояти за себе. У цей період батьки годують їх маленькими шматочками їжі, коли змінюють один одного на посту. Після закінчення цього періоду пташенят годують вже обоє батьків через рівні проміжки часу. Для пошуку корму самець і самка поперемінно вибирають ближні і дальні маршрути, приносячи з собою їжу, що в середньому становить 12% ваги їх власного тіла (близько 600 г). Їжа, якою годують пташенят, може складатися як зі свіжих кальмарів, риби і криля, так і з енергетично багатої легкої олії, яке птахи виробляють у своєму залозистому шлунку[20]. Ця олія, властива більшості буревісникоподібних, виробляється в шлунках з перевареної здобичі і додає птахам характерний запах плісняви.
Пташеням потрібний тривалий час для оперення до того, як вони встануть на крило. Навіть у невеликих видів альбатросів цей період займає 140–170 днів, а у птахів з роду Diomedea він може досягати 280 днів[21]. Як і в інших морських птахів, пташенята альбатросів спочатку набирають вагу більшу, ніж у їхніх батьків, а перед своїм першим польотом використовують ці резерви для розвитку аеродинамічних характеристик тіла, зокрема, для зростання махового пір'я. Кінець кінцем, до моменту свого першого польоту пташенята важать приблизно стільки ж, скільки і їхні батьки. Оперившись і зробивши свій перший самостійний політ, пташеня перестає отримувати допомогу від батьків. Більш того, батьки залишаються байдужими, якщо пташеня покинуло гніздо і відлетіло. На підставі досліджень розсіювання пташенят в океані в перший рік життя, було зроблено припущення про їх природжену міграційну поведінку, що до деякого рівня закладена на генетичному рівні і допомагає їм при визначенні маршрутів міграції[22].
Альбатроси і людина
Походження назви
Сама назва «альбатрос» походить від арабського слова al-câdous або al-ġaţţās (пелікан; буквально «нирець»), яке перейшло до португальської мови у вигляді alcatraz, звідти до англійської мови, де змінилося на albatross, а звідти і до решти європейських мов, включаючи і португальську. Назву Алькатрас також отримав і острів в затоці Сан-Франциско, на якому раніше розташовувалася в'язниця для особливо небезпечних злочинців. Іноді вважають, що португальці першими з європейців познайомилися з альбатросами, коли в 15 столітті обігнули Африку з півдня. Проте Оксфордський словник англійської мови посилається на те, що первинне слово alcatraz відносилося до фрегатів, і лише пізніше під впливом латинського слова albus («білий») трансформувалося в albatross і стало стосуватися світлих альбатросів, а не темних фрегатів[6]. Наукова родова назва Diomedea, дана альбатросам Карлом Ліннеєм, була привласнена на честь давньогрецього героя Діомеда, що брав участь у Троянській війні.
Загрози і охорона
Незважаючи на свою популярність серед моряків, альбатроси не змогли уникнути загроз, прямо або побічно пов'язаних з людською діяльністю. Стародавні полінезійці та індіанці з Алеутських островів полювали на цих птахів, що в деяких випадках привело до зникнення цілих популяцій, як це трапилося на острові Пасхи. Як тільки в Європі стало розвиватися мореплавство, європейці також стали полювати на них заради вживання в їжу або просто для задоволення, ловлячи їх на наживку[23]. Спортивні змагання з лову птахів досягли свого піку під час масового заселення Австралії і припинилися тільки тоді, коли швидкість кораблів стала такою, що з них стало незручно ловити рибу, а також з'явилися закони, що обмежили застосування вогнепальної зброї. У 19 столітті колонії альбатросів на півночі Тихого океану стали відвідувати мисливці за пір'ям для дамських капелюшків, що трохи не привело до вимирання білоспинного альбатроса.
З 21 виду альбатросів, що визнаються Міжнародним союзом охорони природи, 19 знаходяться під охороною цієї організації, оскільки їм так чи інакше загрожує небезпека вимирання, а ще 2 види близькі до цього статусу[24]. Два види — амстердамський (Diomedea amsterdamensis) і чатемський альбатроси (Thalassarche eremita) — визнані такими, що знаходяться в критичному стані. Одним із загрозливих чинників є розвиток рибальства ярусним способом[25], при якому альбатросів й інших морських птахів привертає наживка з рибного тельбуха, насаджена на гачок. Щорічна загибель альбатросів, що попалися на гачок, оцінюється орнітологами від 100 до 300 тис. особин. Посилює ситуацію і неконтрольоване браконьєрське рибальство.

Ще одним чинником ризику є розповсюдження інтродукованих (тобто чужорідних, невластивих даній місцевості, завезених людиною) видів тварин, таких як щурі або здичавілі коти, які нападають на дорослих птахів та поїдають їх яйця і пташенят. Історично альбатроси завжди гніздилися там, де були відсутні наземні хижаки, і, таким чином, у них не виробилося рефлексів захисту. Заподіяти збиток можуть навіть такі дрібні гризуни, як миші: на острові Гоф пташенята тристанського альбатроса були атаковані інтродукованими домовими мишами (Mus musculus), які майже в 300 разів менші їх за розміром[26]. Інтродуковані види можуть заподіяти і непрямий збиток: велика рогата худоба на острові Амстердам спустошила трав'яний покрив, у якому амстердамські альбатроси ховали гнізда.
Серед проблем, з якими зіткнулися багато видів морських птахів, — плаваюче пластмасове сміття. З 1960-х років, коли з'явилися перші пластмасові вироби, кількість сміття в океані істотно зросла. Сміття потрапляє в море найрізноманітнішими способами: з судів, що проходять мимо, з пляжів і з гирл річок. Воно осідає в шлунках птахів, не перетравлюючись, або перекриває шлунково-кишковий тракт так, що птахи страждають від голоду. Дослідження птахів у північній частині Тихого океану показало, що попадання сміття всередину прямо впливає на вагу і фізичний стан птахів[27]. Іноді батьки відригують шматочки пластмаси пташенятам разом з їжею під час годування. Вивчаючи темноспинного альбатроса (Phoebastria immutabilis) на атолі Мідвей, орнітологи виявили у померлих природною смертю пташенят більшу кількість сміття, чим у загиблих у результаті нещасних випадків[28]. Хоча пластмаса безпосередньо не впливає на смертність, вона є причиною фізіологічного стресу і примушує пташенят не відчувати голод, вимагати менше їжі, тим самим зменшуючи свої шанси на виживання.

Вчені і громадські організації, що займаються охороною навколишнього середовища (головним чином, BirdLife International та її партнери), працюють у контакті з урядами країн і риболовецькими компаніями з метою зменшити загрози, з якими стикаються альбатроси. Щоб зменшити смертність птахів при попаданні на гачок при ярусному лові, використовуються додаткові запобіжні засоби: лов у темний час доби, використання наживки на чималій глибині, обваження волосіні й використання пташиних відлякувачів[29]. Наприклад, у результаті сумісного проекту орнітологів і риболовецьких компаній у Новій Зеландії було успішно випробувано пристрій, який опускав волосінь нижче за планку, на яку були в змозі упірнути зникаючі види альбатросів[30].
Вважають, що використання деяких з перерахованих технологій на Фолклендських островах за останні 10 років значно зменшило загибель чорнобривих альбатросів[31]. Крім того, члени природоохоронних організацій працюють над відновленням первинного природного балансу на островах, видаляючи чужорідні види тварин, що загрожують популяції альбатросів.
У результаті зусиль міжнародних природоохоронних організацій у 2001 році було підписано міжнародну Угоду про охорону альбатросів і буревісників (англ. Agreement on the Conservation of Albatrosses and Petrels), яка в 2004 році набрала чинності. Документ зобов'язує сторони, що підписали його, докласти зусиль до зменшення загибелі птахів при вилові риби, скоротити забруднення навколишнього середовища і очистити острови з пташиними колоніями від інтродукованих видів тварин. Станом на 2008 рік угода була ратифікована урядами одинадцяти держав: Австралії, Великої Британії, Іспанії, Нової Зеландії, Перу, Франції, Еквадору, ПАР, Чилі, Аргентини і Норвегії. Крім того, угоду підписала, але ще не ратифікувала Бразилія[32].

Альбатроси в культурі
Альбатросів іноді описують як «найбільш легендарних птахів», з ними пов'язана багато міфів і легенд[21]. Навіть у наші дні багато хто вірить, що спричинення шкоди альбатросові може принести нещастя, хоча це повір'я пішло з відомого твору англійського поета-романтика Семюела Колріджа «Поема про старого моряка», у якій моряка чекає покарання долі за те, що він вбиває альбатроса. Французький поет Шарль Бодлер у вірші «Альбатрос» метафорично порівнює спійманого моряками альбатроса із знедоленим поетом.
У наш час альбатроси часто стають частиною творів поп-культури: наприклад, героями шоу «Albatross» британської комік-групи Монті Пайтон. У фільмі «Місія „Сереніті“» альбатросом називали героїню Саммер Ґлау (англ. Summer Glau), посилаючись на широко розповсюджений метафоричний образ цього птаха.
Систематика
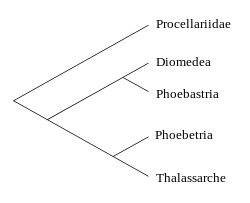
У даний час розрізняють від 13 до 24 видів альбатросів, розбитих на 4 роди. Класифікація на рівні видів знаходиться у стадії обговорення і наукових дебатів, проте більшість учених-орнітологів визнають 21 вид. Серед чотирьох родів (Diomedea, Thalassarche, Phoebastria і Phoebetria) умовно часто виділяють дві морфологічно близькі групи: роди Phoebastria і Diomedea входять до однієї групи, а Thalassarche і Phoebetria — до іншої.
Існує кілька основних гіпотез про положення альбатросів у системі класифікації птахів. Наприклад, у класифікації Сіблі-Алквіста (1990 рік), що ґрунтується на результатах порівняльної гібридизації геномів, всі морські птахи (включаючи й альбатросів), а також і деякі інші групи птахів, поміщені в один величезний ряд лелекоподібних. Орнітологічні товариства Європи, Північної Америки, ПАР, Австралії і Нової Зеландії дотримуються традиційнішої гіпотези, згідно з якою альбатроси є частиною ряду буревісникоподібних. Від інших родин цього ряду вони відрізняються як генетично, так і за морфологічними характеристиками: розміром, будовою ніг і розташуванню носової трубки.
Усередині самої родини історія класифікації таксонів триває вже декілька століть. Спочатку всі альбатроси були зараховані до єдиного роду Diomedea, але в 1852 році німецький ботанік-систематик і зоолог Людвіг Райхенбах розділив їх на чотири роди. Надалі їх об'єднували і знову розділяли кілька разів, внаслідок чого до 1965 року було відомо вже 12 родів альбатросів (проте не більше 8 одночасно): Diomedea, Phoebastria, Thalassarche, Phoebetria, Thalassageron, Diomedella, Nealbutrus, Rhothonia, Julietata, Galapagornis, Laysanornis і Penthirenia.
У 1965 році була зроблена чергова спроба об'єднати всі види птахів у 2 роди: «справжні альбатроси» (Diomedea) і «димчасті альбатроси» (Phoebetria, група птахів, що має риси альбатросів, але в порівнянні з ними «примітивніша»)[33]. Така спрощена система була заснована на морфологічному аналізі Елліота Куеса 1866 року і майже не враховувала пізніші дослідження і навіть ігнорувала деякі припущення самого Куеса.
У 1996 році Ґері Нан (англ. Gary Nunn) з Американського музею природознавства спільно з іншими дослідниками з різних країн почав дослідження мітохондріальної ДНК всіх 14 досліджених на той час видів і прийшов до висновку, що в межах альбатросових можна виділити чотири, а не дві монофілетичні групи[34]. Учені запропонували повернути два з раніше втрачених родів — Phoebastria і Thalassarche, розподіливши частину птахів між ними. Першими такий розподіл визнали Британське орнітологічне товариство (англ. British Ornithologists' Union) і відомства ПАР, а пізніше — і більшість учених у світі.

Якщо відносно класифікації на рівні родів є певний консенсус, то щодо кількості видів існують розбіжності. У минулому різними вченими було описано до 80 різних видів цих птахів, більшість з яких помилково приписувалися молодим птахам інших видів, що значно відрізняються від своїх батьків[35]. Ґрунтуючись на результатах своїх робіт з ідентифікації родів, Кріс Робертсон (англ. Chris Robertson) і Гері Нан у 1998 році запропонували переглянути існуючу систему і виділити 24 види[36] замість 14 відомих на той момент. У цій проміжній системі статус багатьох підвидів був підвищений до рівня виду, що підняло хвилю критики з боку інших орнітологів. Пізніше одні дослідження підтверджували це розділення, а інші його спростовували. У 2004 році на основі результатів аналізу мтДНК і мікросателітів, проведеного Робертсоном і Наном, було досягнуто консенсусу, що види альбатрос антиподів (Diomedea antipodensis) і тристанський альбатрос (Diomedea dabbenena) відмінні від мандрівного альбатроса (Diomedea exulans), проте, з іншого боку, передбачуваний вид Diomedea gibsoni недостатньо відрізняється від альбатроса антиподів[37]. Більшість із запропонованих видів, 21, визнається багатьма вченими групами, і, зокрема, Міжнародним союзом охорони природи, але є і відмінні думки — наприклад, Джон Пенгалларік (англ. John Penhallurick) і Майкл Вінк (англ. Michael Wink) у 2004 році запропонували зменшити кількість видів до 13, включаючи і об'єднання мандрівного альбатроса з амстердамським (Diomedea amsterdamensis)[38]. У будь-якому випадку, орнітологи погоджуються з необхідністю подальших досліджень з метою досягти загального консенсусу.
Види
Види альбатросів за версією Міжнародного союзу охорони природи і організації BirdLife International:
- Альбатрос (Diomedea)
- Альбатрос мандрівний (D. exulans)
- Альбатрос антиподів (D. [exulans] antipodensis)
- Альбатрос острівний (D. [exulans] amsterdamensis)
- Альбатрос тристанський (D. [exulans] dabbenena)
- Альбатрос королівський північний (D. [epomorpha] sanfordi)
- Альбатрос королівський (D. epomophora)
- Phoebastria
- Альбатрос галапагоський (P. irrorata)
- Альбатрос жовтоголовий (P. albatrus)
- Альбатрос чорноногий (P. nigripes)
- Альбатрос гавайський (P. immutabilis)
- Thalassarche
- Альбатрос чорнобровий (T. melanophris )
- Альбатрос кемпбеллівський (T. [melanophris] impavida)
- Альбатрос сірощокий (T. cauta)
- Альбатрос чатемський (T. [cauta] eremita)
- Альбатрос баунтійський (T. [cauta] salvini)
- Альбатрос сіроголовий (T. chrysostoma)
- Альбатрос смугастодзьобий (T. chlororhynchos)
- Thalassarche carteri (T. (chlororhynchos) carteri)
- Альбатрос Буллера (T. bulleri)
- Бурий альбатрос (Phoebetria)
- Альбатрос бурий (P. fusca)
- Альбатрос довгохвостий (P. palpebrata).
Примітки
- Weimerskirch, Henri (Oct 2004). Wherever the Wind May Blow. Natural History Magazine. Прочитане 2011-01-07
- Brooke, M. (2004). Albatrosses And Petrels Across The World Oxford University Press, Oxford, UK ISBN 0-19-850125-0
- Olson, S. L., Hearty, P. J. (2003) «Probable extirpation of а breeding colony of Short-tailed Albatross (Phoebastria albatrus) on Bermuda by Pleistocene sea-level rise» Proceedings of the National Academy of Science 100: (22) 12825-12829. DOI:10.1073/pnas.1934576100
- Lequette, B., Verheyden, C., Jowentin, P. (1989). Olfaction in Subantarctic seabirds: Its phylogenetic and ecological significance. The Condor 91: 732—135.
- Pennycuick, C. J. (1982). «The flight of petrels and albatrosses (Procellariiformes), observed in South Georgia and its vicinity». Philosophical Transactions of the Royal Society of London B 300: 75—106. DOI:10.1098/rstb.1982.0158
- Tickell, W. L. N. (2000). Albatrosses. Sussex: Pica Press, ISBN 1-873403-94-1
- Weimerskirch H, Guionnet T, Martin J, Shaffer SA, Costa DP. (2000) «Fast and fuel efficient? Optimal use of wind by flying albatrosses» Proc Biol Sci 267: (1455) 1869-74. DOI:10.1098/rspb.2000.1223
- Warham, J. (1996). The Behaviour, Population, Biology and Physiology of the Petrels. London: Academic Press, ISBN 0-12-735415-8
- Cocker, M., & Mabey, R., (2005) Birds Britannica London: Chatto & Windus, ISBN 0-7011-6907-9
- Croxall, J. P., Silk, J. R. D., Phillips, R. A., Afanasyev, V., Briggs, D. R., (2005) «Global Circumnaviagtions: Tracking year-round ranges of nonbreeding Albatrosses» Science 307: 249–250. DOI:10.1126/science.1106042
- Croxall, J. P. & Prince, P. A. (1994). «Dead or alive, night or day: how do albatrosses catch squid?» Antarctic Science 6: 155–162
- Prince, P. A., Huin, N., Weimerskirch, H., (1994) «Diving depths of albatrosses» Antarctic Science 6: (3) 353–354.
- Cobley, N. D. (1996). An observation of live prey capture by а Black-browed Albatross Diomedea melanophrys. Marine Ornithology 24: 45–46.
- Fisher, H. I., (1976) Some dynamics of а breeding colony of Laysan Albatrosses. Wilson Bulletin 88: 121–142
- Robertson, C. J. R. (1993). «Survival and longevity of the Northern Royal Albatross Diomedea epomophora sanfordi at Taiaroa Head» 1937-93. Emu 93: 269–276
- Jouventin, P., Monicault, G. de & Blosseville, J.M. (1981) «La danse de l'albatros, Phoebetria fusca». Behaviour 78: 43—80. DOI:10.1163/156853981X00257
- Pickering, S. P. C., & Berrow, S. D. (2001). Courtship behaviour of the Wandering Albatross Diomedea exulans at Bird Island, South Georgia. Marine Ornithology 29: 29—37.
- Anderson, D. J. & Cruz, F. (1998) Biology and management of the Waved Albatross at the Galapagos Islands. Pp. 105–109 in Albatross Biology and Conservation (Roberston, G. & Gales, R. eds) Chipping Norton: Surrey Beatty and & Sons ISBN 0-949324-82-5
- Warham, J. (1990) The Petrels — Their Ecology and Breeding Systems London: Academic Press
- Warham, J. (1976). The incidence, function and ecological significance of petrel stomach oils. Proceedings of the New Zealand Ecological Society 24: 84–93. Архів оригіналу за 24 липня 2006. Процитовано 21 червня 2008.
- Carboneras, C. (1992) «Family Diomedeidae (Albatross)» in Handbook of Birds of the World Vol 1. Barcelona: Lynx Edicions, ISBN 84-87334-10-5
- Åkesson, S., & Weimerskirch, H., (2005) «Albatross Long-Distance Navigation: Comparing Adults And Juveniles» Journal of Navigation 58: 365–373.
- Safina, C. (2002) Eye of the Albatross: Visions of Норі and Survival New York: Henry Holt & Company ISBN 0-8050-6229-7
- IUCN, 2004. Red List: Albatross Species. Retrieved 13 вересня 2005
- Brothers NP. 1991. «Albatross mortality and associated bait loss in the Japanese longline fishery in the southern ocean.» Biological Conservation 55: 255–268.
- BBC News, 2005. Albatross chicks attacked by mice. Retrieved 6 березня 2006.
- Spear, L. B., Ainley, D. G. & Ribic, C. A. (1995). «Incidence of plastic in seabirds from the tropical Pacific, 1984-91: relation with distribution of species, sex, age, season, year and body weight» Marine Environmental Research 40: 123–146
- Auman, H. J., Ludwig, J. P., Giesy, J. P., Colborn, T., (1997) «Plastic ingestion by Laysan Albatross chicks on Sand Island, Midway Atoll, in 1994 and 1995» in Albatross Biology and Conservation (ed by G. Robinson and R. Gales). Surrey Beatty & Sons: Chipping Norton. Pp. 239-44 Архівовано 30 жовтня 2005 у Wayback Machine.
- Food and Agriculture Organisation (1999) The incidental catch of seabirds by longline fisheries: worldwide review and technical guidelines for mitigation. FAO Fisheries Circular No. 937. Food and Agriculture Organization of the United Nations, Rome.
- O'Toole, Decland & Molloy, Janice (2000) «Preliminary performance assessment of an underwater line setting device for pelagic longline fishing» New Zealand Journal of Marine and Freshwater Research 34: 455–461. Архівовано 27 вересня 2007 у Wayback Machine.
- Reid, A. T., Sullivan, B. J., Pompert, J., Enticott, J. W., Black, A. D., (2004) «Seabird mortality associated with Patagonian Toothfish (Dissostichus eleginoides) longliners in Falkland Islands waters.» Emu 104: (4) 317–325
- Australian Antarctic Division Agreement on the Conservation of Albatrosses and Petrels Site Retrieved 2 вересня 2006
- Alexander, W. B., Fleming C. A., Falla R. A., Kuroda N. H., Jouanin C., Rowan M. K., Murphy R. C., Serventy D. L., Salomonsen F., Ticknell W. L. N., Voous K. H., Warham J., Watson G. E., Winterbottom J. M., and Bourne W. R. P. 1965. «Correspondence: The families and genera of the petrels and their names.» Ibis 107: 401-5.
- Nunn, G. B., Cooper, J., Jouventin, P., Robertson, C. J. R. and Robertson G. G. (1996) «Evolutionary relationships among extant albatrosses (Procellariiformes: Diomedeidae) established from complete cytochrome-b gene sequences». Auk 113: 784–801. Архівовано 17 грудня 2008 у Wayback Machine.
- Double, M. C. & Chambers, G. K., (2004). «The need for the parties to the Agreement on Albatrosses and Petrels (ACAP) to establish а robust, defendable and transparent decision-making process for the construction and maintenance of their species lists». Proceedings of the Scientific Meeting of Agreement on Albatrosses and Petrels (ACAP) , Hobart, Australia, 8—9 November 2004
- Robertson, C. J. R. and Nunn, G. B. (1998) «Towards а new taxonomy for albatrosses» in: Proceedings First International Conference on the Biology and Conservation of Albatrosses, G. Robertson & R. Gales (Eds), Chipping Norton: Surrey Beatty & Sons, 13—19
- Burg, T. M., & Croxall, J. P. (2004) «Global population structure and taxonomy of the wandering albatross species complex». Molecular Ecology 13: 2345–2355. Архівовано 27 вересня 2006 у Wayback Machine.
- Penhallurick, J. and Wink, M. (2004). «Analysis of the taxonomy and nomenclature of the Procellariformes bawed on complete nucleotide sequences of the mitochondrial cytochrome b gene» Emu 104: 125–147
Посилання
- Альбатросові / ВУЕ
- Альбатросові / УРЕ
- Альбатросові на сайті zoo-eco.zooclub.ru (рос.)
- Фотографії птахів родини альбатросових — на сайті Animal Diversity Web (англ.)
- Відео альбатросових — на сайті Internet Birds Collection (англ.)
- Save the Albatross — інформація про альбатросів та загрози їм на сайті BirdLife International (англ.)
- Albatrosses of the World на сайті Birds Photo Index — колекція фотографій різних видів (англ.)
- Michael Brooke, John Cox (2004). Albatrosses and Petrels Across the World. ISBN 0-19-850125-0. (англ.)
|
|
Ця стаття належить до вибраних статей Української Вікіпедії. |