Морські птахи
Морські птахи — птахи, пристосовані до життя в морському середовищі. Хоча всі морські птахи в значній мірі відрізняються один від одного як за фізіологією так і поведінкою, вони часто виявляють ознаки конвергентної схожості, що говорить про їх схожу еволюцію в спільних умовах та близьких кормових нішах. Найдавніші морські птахи з'явилися ще в Крейдяний період, а ознак сучасних родин вони набули переважно в епоху Палеогена.

.jpg.webp)
У порівнянні з іншими групами птахів, морські птахи, як правило, живуть довше, розмножуються в зрілішому віці, а пташенят виводять менше, присвячуючи їм значну частину часу. Більшість морських видів гніздяться колоніями, які можуть варіювати від декількох десятків до декількох мільйонів особин. Багато з них щорічно мігрують на значні відстані, перетинаючи екватор, а в деяких випадках навіть здійснюють навколосвітні подорожі. Годуються ці птахи як на поверхні моря, так і на невеликій глибині під водою, а також іноді полюють один одного. Одні види дуже пелагічні, тобто цілий рік проводять на океанічних просторах далеко від берегів, інші ж живуть виключно на узбережжі, а треті певну частину року взагалі проводять далеко від моря.
Морські птахи мають давню спільну історію з людиною: вони служили джерелами їжі для мисливців та вірним орієнтиром для рибалок і мореплавців. Проте в результаті безконтрольної людської діяльності багато видів цих птахів у наш час[коли?] опинилися на межі вимирання, внаслідок чого природоохоронні організації та уряди багатьох країн були змушені розробити спеціальні методи зі збереження біологічної спадщини.
Визначення
Точного визначення того, що якась таксономічна група, наприклад, родина або вид, є морською, у сучасній науковій класифікації не існує, нечисленні запропоновані варіанти такого визначення знаходяться під питанням. За влучним виразом двох орнітологів, «єдиною загальною характеристикою всіх морських птахів є те, що всі вони годуються в солоній воді, але як і з будь-яким твердженням у біології, правду кажучи, деякі цього не роблять»[1]. Проте, існує певний консенсус з приводу того, що усі пінгвіни, буревісникоподібні, пеліканоподібні (за винятком змієшийок) і деяких сивкоподібних (таких як поморники, мартини, крячкові, алькові і водорізи) все ж є морськими. Крім того, до морських птахів часто відносять і плавунців (Phalaropus), які, хоч і вважаються прибережними, але два з трьох видів дев'ять місяців на рік проводять на океанічних просторах, перетинаючи екватор у пошуках їжі.
Гагароподібні і пірникозові, які гніздяться на озерах, а до моря відлітають тільки у зимовий сезон, морськими не вважаються, а зазвичай класифікуються як водоплавні. Не зважаючи на те, що в родині качкових (Anatidae) ціла підродина крехових (Merginae) також зимує в морі, за загальною згодою ці птахи також зазвичай не включаються в список морських видів. Багато болотяних птахів, зокрема чаплі, також значну частину часу проводять на морських просторах, мешкаючи поблизу від узбережжя, але й вони на загальну думку морськими не є.
Еволюція
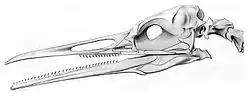
Завдяки літологічному осадковому середовищу (тобто морській акваторії, де тверді частинки легко осідають на дно) збереглася велика кількість викопних залишків морських птахів[2]. Найбільш ранні знахідки відносяться до Крейдяного періоду — майже повні скелети представників стародавнього ряду гесперорнісоподібних (Hesperornithiformes) були виявлені в американському штаті Канзас. Ці зубаті нелітаючі птахи, зокрема схожий на сучасних гагар вид Hesperornis regalis, непогано плавали і упірнали подібно до пірникоз і гагар[3][4]. Вважається, що рід Hesperornis виявився тупиковою гілкою еволюції і не залишив після себе нащадків, проте і предки сучасних морських птахів також беруть свій початок у Крейдяному періоді. Серед таких стародавніх видів можна назвати Tytthostonyx glauconiticus, що можливо відноситься до буревісникоподібних. Пізніше, в епоху Палеогена, на океанських просторах домінували ранні буревісники і гігантські пінгвіни, а також представники двох вимерлих родин — Pelagornithidae, у яких розмах крил досягав 5 м, і Plotopteridae (група крупних морських птахів, схожих на пінгвінів)[5]. Розквіт сучасних родів почався в період Міоцена, а рід справжніх буревісників (куди зокрема входять звичайний (Puffinus puffinus) і сірий (Puffinus griseus) буревісники) з'явився ще раніше, в Олігоцені[2]. Здається, найбільше біорізноманіття морських птахів спостерігалося в пізньому Міоцені і Пліоцені. У кінці останнього Харчовий ланцюжок (система харчових зв'язків — поїдання одних організмів іншими) зазнав значних змін, зважаючи на вимирання значної кількості морських мешканців, і кінець кінцем збільшення числа морських ссавців привело до зменшення різноманіття морських птахів[6].
Опис
Адаптація до морських умов
У морських птахів спостерігається ряд морфологічних особливостей, що дозволяють їм виживати в морі. По будові крил можна сказати, яку екологічну нішу займали даний конкретний вид або родина протягом своєї еволюції, зокрема, дивлячись на форму крила або навантаження на нього, учений може визначити кормову поведінку птаха. Наприклад, довгі крила разом з малим питомим навантаженням властиві пелагічним видам (тобто повітряним видам, що мешкають далеко від берегів), а короткі крила характерні для видів, що активно упірнають[7]. Такі птахи, як мандрівний альбатрос (Diomedea exulans), який здатний долати величезні морські відстані у пошуках їжі, багато в чому втратили здібність до активного польоту і безпосередньо залежать від динамічного ширяння (при якому енергія отримується від переміщення в горизонтальних повітряних потоках з різною швидкістю вітру) або похилого ширяння (при якому енергія отримується від постійної зміни шарів повітря із різними швидкостями)[8]. Практично у всіх морських птахів лапи перетинкові, що допомагає їм при пересуванні по воді, а в деяких випадках і при пірнанні. У буревісникоподібних, на відміну від більшості інших птахів, дуже добре розвинені органи нюху, що дозволяє їм знаходити джерела їжі, сильно розкидані на безкрайніх просторах океану[9], і, можливо, вказати місцезнаходження косяків риб.
Сольові залози, розташовані в голові в області носової порожнини, використовуються для виділення концентрованих розчинів хлористого натрію, що споживається разом з водою і їжею, а також для регуляції осмотичного тиску крові і внутріклітинної рідини[10].

За винятком бакланових і деяких крячків, у всіх морських птахів оперення не намокає. Хоча водозахисне оперення часто зустрічається і в інших птахів, що ведуть наземний спосіб життя, у морських видів кількість такого пір'я незрівнянно більша. Таке густе оперення чудово захищає їх від намокання, а товстий шар пуху зігріває тіло. У бакланів же, чудових нирців за рибою, пір'я формує унікальний у своєму роді шар, який хоч і намокає, але все-таки порівняно з іншими видами зберігає тонкий повітряний прошарок[11]. Така особливість збільшує їх питому вагу і дозволяє довше знаходитися під водою, проте також і зберігає тепло тіла при контакті з водою. В порівнянні з іншими птахами у більшості морських видів оперення менш барвисте і, як правило, являє собою різні варіації чорного, білого і сірого кольорів[7].
Лише у небагатьох тропічних видів і деяких пінгвінів забарвлення більш-менш різноколірне, у інших же колір виявляється тільки на дзьобі і лапах. Вважається, що у багатьох випадках оперення має захисне забарвлення (наприклад, оперення антарктичного пріона (Pachyptila desolata) подібне до забарвлення військових кораблів та зливає його з поверхнею моря[7]), також білі груди багатьох хижих птахів приховують їх від потенційних жертв.
Харчування
В процесі еволюції птахи навчилися здобувати собі їжу в морях і океанах, що неодмінно відбилося на їх фізіології і поведінці. Використовуючи схожі стратегії при здобичі корму і близькі адаптації при вирішенні загальних проблем, різні родини і роди отримали конвергентну схожість, тобто загальні ознаки в результаті конвергентної еволюції. Прикладом такої еволюції можна назвати схожих один алькових і пінгвинів, що відносяться до різних родин. В цілому вчені розрізняють чотири основні стратегії при здобичі корму в морському середовищі: здобич корму на поверхні, пірнання на глибину з повітря, підводне полювання за допомогою переслідування і полювання за вищими хребетними тваринами. У свою чергу кожна з цих стратегій розділяється на ряд варіацій.
Добування корму на поверхні
Багато морських птахів шукають собі прожиток винятково на водній поверхні завдяки морським течіям, які виштовхують Криль, рибу, кальмарів та інших морських мешканців на невелику глибину, де птахові досить опустити під воду голову.

Поверхневе полювання розділяється на два підходи: полювання на льоту (наприклад, у тайфунників (Pterodroma), фрегатів (Fregata) і качурок (Hydrobatidae)) і полювання на плаву (як це практикують фульмари (Fulmarus), мартини і багато буревісників). Полювання на льоту вимагає особливої спритності, яке полягає або в спробі схопити ласощі з льоту (як у фрегатів), або в зависанні над водою (як у деяких альків) [12]. Деякі з них навіть не опускаються на поверхню води, а наприклад фрегати насилу можуть злетіти знову, якщо опиняються на поверхні[13]. Ще однією родиною птахів, які не приводнюються при полюванні, є водорізові. Їхня тактика полягає в тому, що вони летять близько над поверхнею, опустивши у воду своє піддзьоб'я. Як тільки дзьоб натрапляє на що-небудь тверде, він вмить закривається. Ця незвичайна особливість водорізів відбилася на їх морфології — піддзьоб'я у них довше за наддзьоб'я.
Унікальна будова дзьоба часто спостерігається також у птахів, що полюють на плаву. У пріонів (Pachyptila) дзьоб плоский і має спеціальні пластинки-фільтри, що називаються ламелами і виціджують планктон[14]. У багатьох альбатросів та буревісників дзьоб загнутий на кінці, що допомагає ним ловити жертву, що швидко рухається. У мартинів, діапазон харчування яких досить широкий, дзьоб має більш-менш загальну структуру.
Підводне полювання

Підводне полювання створює сильніший тиск (як еволюційний, так і фізіологічний), проте в нагороду значно збільшує кормову базу на обмеженій території. Поступальний рух під водою може виконаються як за допомогою крил (наприклад, у пінгвінів, алькових і пірнаючих буревісників (Pelecanoididae)), так і за допомогою ніг (наприклад, у бакланових, пірникоз, гагароподібних і деяких качок, що харчуються рибою). Як правило, упірнання за допомогою крил дозволяє рухатися швидше, ніж за допомогою ніг[2]. Проте в обох випадках використання крил або ніг при русі під водою обмежує їх можливості в інших ситуаціях: гагароподібні й пірникози по землі якщо й пересуваються, то насилу; пінгвіни не вміють літати, а алькові хоч і літають, але роблять ці маломаневрово і незграбно. Наприклад, для гагарки (Alca torda) з родини алькових енергії для польоту потрібно на 64 % більше, ніж для такого ж розміру буревістника[15]. Багато ж буревісникоподібних володіють проміжними характеристиками: в порівнянні з типовими птахами, що упірнають за допомогою крил, їх крила довші, проте навантаження на крило вище, ніж у птахів, що полюють на поверхні. Це дозволяє як упірнати на чималу глибину, так і подорожувати на значні відстані. Мабуть, найвразливішою виглядає тактика підводного полювання у тонкодзьобого буревісника (Puffinus tenuirostris), здатного у пошуках їжі опуститися на глибину до 70 м[16]. Деяка здатність пірнання існує і у деяких альбатросів: наприклад, димчасті альбатроси (Phoebetria) занурюються на глибину до 12 м[17]. Зі всіх морських птахів, що полюють під водою за допомогою крил, в повітрі найефективніші альбатроси, і невипадково серед пірнаючих птахів вони займають останнє місце. Підводні хижаки домінують в полярних і приполярних водах, а в теплішому кліматі вони енергетично неефективні. Маючи обмежені здібності до польоту, ці птахи не мають можливості до здобичі прожитку на чималій території, і це особливо гостро відчувається в сезон розмноження, коли голодні пташенята вимагають повноцінного харчування.
Пірнання
Олуші (Sula), фаетони (Phaethon), деякі крячкові і бурі пелікани полюють за жертвою, що швидко рухається, упірнаючи у воду з літу. При цьому опір силі виштовхування (збільшеній через велику кількість бульбашок повітря між пір'ям) забезпечується енергією, накопиченою під час польоту[18]. Таким чином, пірнаючі птахи витрачають менше енергії в порівнянні з підводними мисливцями і менш обмежені певною територією, що привело їх до більшого розповсюдження, зокрема в тропіках. В цілому пірнання вважається найбільш адаптованою тактикою добування корму у морських птахів. Інші птахи, що не спеціалізуються на такому способі здобування прожитку (як мартини і поморники) також при нагоді здатні пірнати на льоту, але роблять це менш ефективно і з меншої висоти. У бурих пеліканів навичка занурення виробляється тільки через декілька років; дорослі птахи пірнають з висоти 20 м над водою і перед ударом об воду підводять тулуб, щоб уникнути пошкоджень[19]. Вважається, що для пірнаючих птахів необхідна достатньо чиста прозора вода для ефективного огляду з повітря[20], проте при їх домінуванні в тропіках зв'язок між пірнанням і чистою водою виглядає непереконливо[21]. Деякі пірнаючі птахи (також як і деякі мисливці на поверхні) залежать від дельфінів і тунців, які виштовхують косяки риб ближче до поверхні моря[22].
Клептопаразитизм, харчування падаллю і хижацтво
Дана всеосяжна категорія птахів відноситься до ще однієї стратегії добування прожитку, яка піднімається на наступний трофічний рівень. Клептопаразитами називають таких птахів (і не тільки птахів), чия стратегія полягає у відбиранні здобичі у інших птахів. Такий стиль поведінки найхарактерніший для фрегатів і поморників, хоча мартини, крячки та інші види морських птахів також при нагоді використовують дану практику[23]. Нічна активність деяких морських птахів під час гніздування часто пояснюють захистом від зростаючої активності хижаків в світлий час доби[24]. Вважається, що клептопаразитизм не грає ключової ролі в поведінці будь-якого виду, а є лише додатковим способом добування корму разом з полюванням[2]. Дослідження, проведене орнітологами у відношенні великого фрегата (Fregata minor), що краде їжу у маскової олуші (Sula dactylatra), показало, що фрегати за рахунок клептопаразитизму здатні до 40 % забезпечити себе прожитком, але в середньому ця цифра становить тільки 5 %[25]. Багато видів мартинів та інших морських птахів, як північний гігантський буревісник (Macronectes halli) при нагоді харчуються падаллю — іншими загиблими птахами або ссавцями. Падальщиками також є деякі види альбатросів — аналіз проковтнутих ними кальмарів показав, що деякі з них були дуже великими, щоб бути спійманими живцем. Крім того, в шлунках альбатросів опинилися види, які швидше за все були їм недоступні зважаючи на різний ареал мешкання[26]. Деякі морські птахи, такі як мартини, поморники і південні гігантські буревісники (Macronectes giganteus) полюють за яйцями, пташенятами і навіть за невеликими особинами інших морських птахів в районах великих колоній[27].
Життєвий цикл

Життєвий цикл морських видів значно відрізняється від птахів, що живуть на материку. Згідно з теорією r/K-відбору (наукова теорія, що представляє два типи стратегій популяцій: r-стратегія «прагне» до максимального використання ресурсів речовини і енергії за рахунок високої швидкості росту чисельності, а K-стратегія зазвичай знаходиться в стані рівноваги із своїми ресурсами[28][29]) морські птахи прихильні K-стратегії, живуть набагато довше (в цілому 12-60 років), плодитися починають пізніше (до 10 років) і у виховання потомства вкладають більше сил[2][30]. У більшості видів кладка яєць відбувається тільки один раз на рік у випадку, якщо до цього кладка не закінчилася невдачею (бувають і винятки, як у алеутської конюги (Ptychoramphus aleuticus)) [31], а також у багатьох випадках щорічно відкладається тільки одне яйце (наприклад, у буревісникоподібних і олушевих)[14].
Процес вигодовування і виховання пташенят — один з найтриваліших серед всіх груп птахів і може продовжуватися до шести місяців. Наприклад, пташенята тонкодзьобої кайри (Uria aalge), що вже встали на крило, ще декілька місяців проводять в морі разом зі своїм батьком[15]. У фрегатів турбота про потомство займає рекордний серед всіх птахів час: пташенята починають літати тільки через 4-6 місяців, і ще протягом 14 місяців продовжують отримувати допомогу від батьків[32]. У деяких видів такий тривалий процес розмноження привів до того, що птахи починають гніздитися тільки раз на два роки. Такий життєвий цикл швидше за все сформувався завдяки специфічним умовам життя в морі: розсіяній кормовій базі, частим невдалим спробам відтворити потомство внаслідок погодних умов і відносно малій (у порівнянні з наземними видами) кількості хижаків[2].
У всіх морських видів птахів, за винятком плавунців, обоє батьки доглядають за потомством, а пари зазвичай сезонно моногамні. Це відбувається тому, що пташенята вимагають великої уваги, а пошуки їжі можуть відбуватися на значній відстані від гнізда. У мартинів, алькових, пінгвінів та інших морських птахів пари зберігаються протягом декількох сезонів, а у багатьох буревісникоподібних — протягом всього життя[14]. У альбатросів і буревісників, чиї пари зберігаються на все життя, процес залицяння може тривати декілька років, перш ніж пара остаточно сформується і породить потомство. У альбатросів залицяння супроводжується характерними шлюбними танцями [33].
Розмноження і колонії

95 % всіх морських птахів розмножуються в колоніях, розмір яких може досягати рекордних серед птахів величин і бути вражаючим природним видовищем. Колонії, що включають понад мільйон птахів, зустрічаються як на широті тропіків (наприклад, в районі острова Кіритіматі в Тихому океані), так і за полярним колом (як в Антарктиці, так і в Арктиці). У колонії птахи збираються виключно з метою розмноження, решту часу вони можуть скупчуватися у великій кількості тільки в місцях великої концентрації корму.
Самі по собі колонії значно відрізняються одна від одної. Щільність гніздовій буває як досить низькою (як у альбатросів), так і дуже високою (як у кайр і чистунів). Частіше всього в межах однієї колонії гніздяться кілька видів птахів, що іноді займають різні екологічні ніші. Гнізда можуть влаштовуватися на деревах (у разі їх наявності), на землі (іноді гнізда як такого може і не бути), на скелях, в скелястих печерах або земляних норах. Конкуренція за місце може бути дуже високою як серед птахів одного виду, так і серед різних видів. Наприклад, агресивніший темний крячок (Onychoprion fuscata) може витіснити інших птахів з найзручніших насиджених місць[34]. Щоб зменшити конкуренцію агресивнішим клинохвостим буревісникам (Puffinus pacificus), тропічний бонійський тайфунник (Pterodroma hypoleuca) надає перевагу гніздуванню в зимову пору року. Як тільки починається літній сезон розмноження у клинохвостого буревісника, він знищує пташенят тайфунника і займає його нори[35].
Багато морських птахів демонструють дивовижну прихильність до одного й того ж місця, щорічно повертаючись до тієї ж самої нори, гнізда або району протягом багатьох років; вони рішуче охороняють це місце від суперників[2]. Така поведінка покращує народжуваність, дає можливість одним і тим же самцеві і самці спаровуватися знову і скорочує час на пошуки нового місця для гнізда[36]. Птахи, що тільки досягли статевої зрілості, також прагнуть влаштувати своє перше гніздо недалеко від місця, де вилупилися самі. Така тенденція, відома як філопатрія, досить сильно розвинена у багатьох видів: наприклад, автори дослідження, проведеного відносно темноспинного альбатроса (Phoebastria immutabilis), прийшли до висновку, що відстань між гніздом пташеняти і його першим власним гніздом в середньому становить 22 м[37]. Інше дослідження, цього разу проведене в районі Корсики у відношенні строкатого атлантичного буревісника (Calonectris diomedea), з'ясувало що 9 з 61 пташеняти чоловічої статі, що повернулося в свою рідну колонію, відтворили своє перше потомство в тій же самій норі, де і вилупилися самі, а двоє з них спарувалися зі своєю власною матір'ю[38].
Як правило, колонії влаштовуються на островах, скелях або у високогір'ях, недоступних для наземних хижих ссавців[39]. Вважається, що це дає додатковий захист морським птахам, які на землі часто поводяться досить незграбно. У загальному випадку колонії взагалі часто створюються видами, які не охороняють свою власну кормову територію (наприклад, у серпокрильцевих, що мають широкий кормовий діапазон), — ймовірно тому така поведінка дуже часто зустрічається у морських птахів. Ще однією можливою перевагою колоній є те, що вони виступають в ролі інформаційного центру, коли птахи, що прямують до моря в пошуках корму, можуть визначити місцезнаходження косяків риб та іншої кормової бази по напрямку польоту птахів того ж виду, що повертаються із здобиччю. У колоній є і свої недоліки: вони можуть стати джерелом масової епідемії або привернути увагу хижаків, головним чином інших птахів[40].
Міграція

Часто морські птахи, як і птахи взагалі, щорічно здійснюють сезонні міграції. Абсолютним рекордсменом за дальністю перельоту є полярний крячок (Sterna paradisaea), який літом гніздиться в арктичній і субарктичній зоні північної півкулі, після чого перетинає екватор і зимує в районі Антарктиди. Інші види також здійснюють трансекваторіальні подорожі, як з півночі на південь, так і з півдня на північ. Популяція елегантного крячка (Thalasseus elegans) в районі Нижньої Каліфорнії в Мексиці після закінчення сезону розмноження роздвоюється: частина птахів вилітає на північ до району узбережжя центральної Каліфорнії в США, а інша частина летить на південь до берегів Перу і Чилі, де годується у водах Перуанскої океанської течії[41]. Сірий буревісник (Puffinus griseus) за рік здійснює шлях близько 64 тис. км; він гніздиться в районі Нової Зеландії і Чилі, а решту часу проводить на півночі Тихого океану в районі Аляски, Японії і північної Каліфорнії[42].
Інші перелітні птахи здійснюють коротші подорожі, їх розповсюдження в океані в основному залежить від наявності джерел харчування. Якщо в якому-небудь регіоні умови невідповідні, птахи переміщаються в райони із сприятливішими умовами, а молоді особини іноді роблять це постійно[43]. Тільки вставши на крило, пташенята, що подорослішали, часто розсіваються далі за дорослі особини, а діапазон їх розповсюдження часто ширший. У деяких птахів, як у алькових, певного району зимування не існує, але вони кочують на південь з приходом холодів[15]. Інші види, такі як качуркові (Hydrobatinae), пірнаючі буревісники (Pelecanoides) і бакланові (Phalacrocoracidae) не мігрують зовсім, залишаючись поблизу від колоній цілий рік.
На суші
Хоча визначення морських птахів припускає, що вони проводять своє життя в океані, в багатьох родинах є види, які частину свого річного циклу, іноді дуже велику, проводять в стороні від моря. Більш того, багато видів гніздяться в десятках, сотнях і навіть тисячах кілометрах від узбережжя. Деякі з цих птахів все ж таки вилітають у пошуках корму до моря: наприклад, сніжний буревісник (Pagodroma nivea), гнізда якого знаходять за 500 кілометрів від берега в Антарктиді, просто не в змозі знайти собі прожиток в цьому районі[44]. Азіатський довгодзьобий пижик (Brachyramphus marmoratus) свої гнізда будує в перестійному лісі на гілках великих хвойних дерев[45]. Каліфорнійський мартин (Larus californicus) гніздиться на внутрішніх озерах, а в зимовий час вилітає до узбережжя[46]. У деяких видів бакланових, пеліканів, мартинів і крячкових трапляються окремі особини, які взагалі ніколи не бувають в морі, а своє життя проводять на озерах, річках, болотах і, у разі мартинів, в містах і на сільськогосподарських угіддях. У таких випадках говорять, що ці наземні і прісноводі птахи еволюціонували від своїх морських предків[7]. У деяких морських птахів, головним чином поморників і плавунців, що гніздяться в тундрі, частина міграції відбувається над сушею.
Деякі типово морські види, такі як буревісникові, алькові і олушеві, часом випадково залітають углиб материків. Найчастіше це трапляється з молодими недосвідченими птахами, але іноді у разі крупного шторму велика кількість птахів, що терплять лихо, шукає собі укриття на суші[47].
Взаємодія з людиною
Морські птахи і рибальство
Морські птахи довгий час асоціювалися з мореплавацями і рибаками, і обидві ці групи людей стикалися з перевагами і недоліками такого сусідства.
Рибаки традиційно використовували птахів як орієнтир при пошуку косяків риб[22], мілин з рибними ресурсами і потенційних підходів до берега. Відомо, що стародавні полінезійці використовували відомий зв'язок морських птахів з берегом для знаходження крихітних ділянок суші в Тихому океані[2]. Далеко від дому рибаки харчувалися птахами і використовували їх як наживку. Більш того, прив'язані за мотузок баклани інколи безпосередньо використовувалися для рибного лову. Гуано (послід, що розклався природним чином) з пташиних колоній використовувався рибаками як Добриво для довколишніх ділянок моря.
Серед негативних ефектів для рибальства головним чином є гальмування розвитку аквакультури (культивування водних організмів), оскільки штучно зібрана в одному місці риба неодмінно привертала птахів[48]. Крім того, рибаки, що ловлять рибу на гачок, стикалися з крадіжкою наживки. Є і докази того, що в деяких випадках морські птахи сприяли виснаженню морських запасів, проте ефект від такого збитку виглядає значно меншим, ніж аналогічний збиток від морських ссавців або хижих риб, таких як тунець[2].

З іншого боку, деякі види морських птахів також отримують перевагу від взаємодії з рибаками, зокрема споживаючи в їжу викинуту рибу і потрухи. Наприклад, такі покидьки складають до 30 % всього раціону птахів в Північному морі, а для деяких популяцій ця цифра зростає до 70 %[49]. Ще одним ефектом такої взаємодії може стати зміна чисельності птахів: наприклад, розповсюдження кочівних буревісників (Fulmaris glacialis) на Британських островах приписують саме розвиненому рибальству[50]. Від наявності покидьків більше всього переваг отримують птахи, що харчуються на поверхні моря, такі як олуші і буревісники; а для підводних мисливців, таких як пінгвіни, вони можуть заподіяти лише збиток.
Рибальство саме по собі може стати джерелом загроз для деяких видів птахів, зокрема для альбатросів, що довго живуть і поволі розмножуються, і ця проблема викликає зростаючий інтерес екологів. Окрім втрати кормової бази при надмірному лові риби, ці птахи можуть зіткнутися із загибеллю в риболовецьких снастях: наприклад, щорічно близько 100 тис. альбатросів попадаються на гачок з наживкою при ярусному лові на тунця[51][52] або гинуть в дрифтерних мережах[53]. У загальному масштабі в результаті схожих обставин щороку гинуть сотні тисяч птахів, і особливу турботу викликають рідкісні види (наприклад, чисельність короткохвостих альбатросів (Phoebastria albatrus) у світі скоротилася до лише близько 1000 особин).
Використання
Полювання на птахів і вживання в їжу їх яєць сприяли значному зниженню чисельності популяцій багатьох видів, а в деяких випадках, як це відбулося з великою гагаркою (Pinguinus impennis) і стеллеровим бакланом (Phalacrocorax perspicillatus), до їх повного вимирання. Випадки полювання людей, що живуть на узбережжі, на морських птахів були відомі у всі часи; найбільш раннє свідоцтво, що дійшло до нас, відноситься до періоду приблизно 5000 років до н. е.: археологічні розкопки горбів, утворених з покидьків первісної людини на півдні Чилі, показали останки альбатросів, бакланів і буревесників[54]. Скорочення природного біорізноманіття відбувалося в багатьох регіонах планети: наприклад, принаймні 20 видів птахів з 29, що колись гніздилися в районі острова Пасхи, зникли їх цих місць. У 19 столітті полювання на морських птахів з метою отримання жиру і пір'я для дамських капелюшків досягло промислових масштабів. Однєю з великих галузей промисловості в Новій Зеландії і Тасманії стало збирання пташенят буревісників[55].
На Фолклендських островах після їх освоєння люди стали знищувати сотні тисяч пінгвінів на рік з метою отримання жиру. Довгий час джерелом прожитку для дальніх мореплавців служили пташині яйця. Яйцями також харчувалися і мешканці узбережжя, недалеко від якого існували пташині колонії: наприклад, мисливці за яйцями з Сан-Франциско в середині 19 століття щорічно здобували до пів-мільйона яєць з островів Фараллон — з того часу чисельність птахів в цьому районі ще остаточно не відновилася[56].
Полювання на птахів і за їх яйцями продовжується і сьогодні, хоч і не в таких масштабах як раніше, і під частковим контролем. Наприклад, аборигени племені маорі з острова Стюарт поблизу Нової Зеландії, як і століття тому, продовжують полювати за пташенятами сірого буревісника (Puffinus griseus) своїм традиційним способом, відомим як каїтіакітанга (англ. kaitiakitanga) ради отримання доброго врожаю. Проте останніми роками спільно із співробітниками Університету Отаго (англ. University of Otago) проводяться дослідження популяції цих птахів. Іншим прикладом може служити Гренландія, де безконтрольне полювання за морськими птахами привело до різкого скорочення популяції багатьох видів[57].
Інші загрози

Окрім власне знищення морських птахів, інші людські чинники також приводять до зменшення чисельності і навіть вимирання багатьох видів, колоній і популяцій. Серед таких чинників найбільшу небезпеку представляють інтродуковані види (тобто види, нехарактерні для даної території, так або інакше завезені туди людиною). Морські птахи, що живуть на маленьких островах, багато в чому втратили свою здатність захищатися від хижаків[39]. Здичавілі кішки полюють на птахів і здатні зловити жертву розміром з альбатроса, багато інтродукованих гризунів, такі як малий щур (Rattus exulans), поїдають яйця птахів, заховані в норах. Кози, велика рогата худоба, зайці та інші травоїдні тварини поїдають рослинність, в якій ховаються пташині гнізда[58]. Причиною неспокою можуть стати навіть доброчесні туристи, які, зполохуючи з колонії зграї птахів, залишають беззахисними пташенят на поживу хижакам.
Неспокій також викликають токсичні речовини і забруднення навколишнього середовища. Як вищі хижаки, багато видів раніше страждали від руйнівної дії ДДТ, поки його застосування не було заборонене. Крім того, ДДТ викликав проблеми ембріонального розвитку і спотворив співвідношення між статями у західного мартина (Larus occidentalis) в південній Каліфорнії[59]. Істотну загрозу для морських птахів представляє витік нафтопродуктів з танкерів: нафта як сама по собі є токсичною, так і просочує пір'я птахів, внаслідок чого ті втрачають водонепроникність[60]. Забруднення нафтою в основному загрожує видам з обмеженим ареалом і популяціям, що вже ослабіли.
Охорона
Загрози, з якими зіткнулися морські птахи, не залишили без уваги учених-орнітологів і екологічні організації. Ще в 1903 році президент США Теодор Рузвельт виступив з пропозицією оголосити Острів Пелікан біля узбережжя Флориди національним заповідником з метою збереження пташиних колоній, у тому числі і колоній бурого пелікана (Pelecanus occidentalis)[61], а в 1909 році встав на захист островів Фараллон біля берегів Каліфорнії. Сьогодні багато пташиних колоній, наприклад на островах Херон (англ. Heron Island) в Австралії і Тріангл (англ. Triangle Island) в Британській Колумбії так чи інакше знаходяться під охороною національних законодавств.
В результаті спеціальних проектів по відновленню природної спадщини на островах поступово виводяться інтродуковані види, що витісняють рідну флору і фауну. Наприклад, острів Вознесіння в Атлантичному океані був повністю очищений від здичавілих кішок, з Алеутських островів були витиснені песці[62], на островах Кемпбелл в Тихому океані знищені щури. В результаті цих заходів багато видів птахів, що раніше знаходилися під загрозою, а в деяких випадках і зниклі з певних територій, збільшили чисельність своїх популяцій. На острові Вознесення птахи почали гніздитися знову через сотні років[63].
Смертність птахів в результаті попадання в мережі може бути значно знижена за допомогою використання більш щадних методів рибальства, такими як використання наживки в нічний час і на глибині, збільшення вантажу на волосінях і використання спеціальних відлякувачів[64]. Міжнародна заборона на використання дрифтерних риболовних мереж також сприяє зменшенню смертності серед птахів та інших мешканців моря.
Одним з великих британських проектів стало заснування в 2000 році Шотландського центра морських птахів (англ. Scottish Seabird Centre) у місті Норт-Бервік. Для відвідувачів пропонуються відеопрограми, що розповідають про птахів, про їх загрози і заходи по оберіганню та пряме відео, зняте відеокамерами на островах Басс-Рок, Фідра і ряді інших. Ці території є рідним домом величезних колоній олуш, тупиків (Fratercula), поморників та інших птахів. Доходи від туризму прямують як для підтримки місцевих общин, так і на природоохоронні потреби. Інші місця гніздування морських птахів також привертають багато туристів. Відомо, що наприклад колонію південного королівського альбатроса (Diomedea sanfordi) на мисі Таяроа (Taiaroa Head) в Новій Зеландії відвідують близько 40 тис. туристів на рік[14].
Численні громадські організації, такі як BirdLife International і Королівське товариство охорони птахів (англ. Royal Society for the Protection of Birds), звернули увагу рибалок на важке становище альбатросів та інших крупних морських птахів, в яке вони потрапили у зв'язку з побічною смертністю птахів при використанні деяких методів лову риби. В результаті цих зусиль було підписано міжнародну Угоду про охорону альбатросів і буревісників (англ. Agreement on the Conservation of Albatrosses and Petrels), яку станом на 2008 рік ратифікували 11 держав (Австралія, Велика Британія, Іспанія, Нова Зеландія, Перу, Франція, Еквадор, ПАР, Чилі, Аргентина і Норвегія)[65].
Морські птахи в культурі

Більшість морських птахів недостатньо вивчені або про них майже нічого невідомо, оскільки вони мешкають далеко в океані і гніздяться на ізольованих скелях. Проте деякі птахи, такі як альбатроси, буревісники і мартини, стали в народі дуже популярними. Зокрема, альбатросів називають «найлегендарнішим птахом»[66]: з ними пов'язана множина міфів і легенд. Навіть в наші дні багато хто вірить, що спричинення шкоди альбатросові може принести нещастя, хоча це повір'я пішло з відомої поеми англійського поета-романтика Семюела Колріджа «Поема про старого моряка», у якій моряка чекає покарання долі за те, що він вбиває альбатроса і вішає його труп собі на шию.
Найближче люди знайомі з мартинами, які давно міцно влаштувалися в густонаселених містах і на сміттєвих звалищах. Крім того, людям добре знайомий їх небоязливий характер. З цих причин ці птахи міцно увійшли до культури багатьох народів: їх зображають як метафору (як наприклад в повісті-притчі «Мартин на ім'я Джонатан Лівінгстон» Річарда Баха) або щоб показати близькість моря (наприклад, в романі Володар перснів Джона Толкіна мартини зображені на знаках Гондора).
Серед інших птахів можна відзначити пеліканів, які довгий час асоціювалися з альтруїзмом завдяки міфу західних християн, що за умовами голоду пелікани годують пташенят власною кров'ю.
Систематика
Такі групи птахів зазвичай класифікують як морські (родини і підродини списку):
Пінгвіноподібні (Sphenisciformes) — Антарктида і моря південної півкулі. 16 видів
- Пінгвінові (Spheniscidae)
Буревісникоподібні (Procellariiformes) — Пелагічні види всіх океанів. 93 види
- Альбатросові (Diomedeidae)
- Буревісникові (Procellariidae)
- Буревісникові (Procellariinae)
- Качуркові (Hydrobatinae)
- Пірнаючі буревісники (Pelecanoididae)
Пеліканоподібні (Pelecaniformes) — По всьому світу. 57 видів
- Пеліканові (Pelecanidae)
- Олушеві (Sulidae)
- Бакланові (Phalacrocoracidae)
- Фрегатові (Fregatidae)
- Фаетонові (Phaethontidae)
Сивкоподібні (Charadriiformes) — По всьому світу. 305 видів
- Поморникові (Stercorariidae)
- Мартинові (Laridae)
- Мартинові (Larinae)
- Крячкові (Sterninae)
- Водорізові (Rhynchopinae)
- Алькові (Alcidae)
Посилання
- (англ. "The one common characteristic that all seabirds share is that they feed in saltwater; but, as seems to be true with any statement in biology, some do not") Schreiber, Elizabeth A. & Burger, Joanne.(2001.) Biology of Marine Birds, Boca Raton:CRC Press, ISBN 0-8493-9882-7
- Schreiber, Elizabeth A. & Burger, Joanne (2001). Biology of Marine Birds. Boca Raton: CRC Press. ISBN 0-8493-9882-7.
- Johansson LC, Lindhe Norberg UM. (2001). Lift-based paddling in diving grebe. J Exp Biol. 204 (10): 1687–1696. Процитовано 7 січня 2011.
- Gregory, J. (1952). The Jaws of the Cretaceous Toothed Birds, Ichthyornis and Hesperornis. Condor 54 (2): 73–88. Архів оригіналу за 6 червня 2011. Процитовано 31 травня 2008.
- Goedert, J. (1989). Giant Late Eocene Marine Birds (Pelecaniformes: Pelagornithidae) from Northwestern Oregon. Journal of Paleontology 63 (6): 939—944.
- Olson, S. & Hasegawa, Y. (1979). Fossil Counterparts of Giant Penguins from the North Pacific. Science 206 (4419): 688—689.
- Gaston, Anthony J. (2004). Seabirds: A Natural History. New Haven: Yale University Press. ISBN 0-300-10406-5.
- Pennycuick, C. J. (1982). «The flight of petrels and albatrosses (Procellariiformes), observed in South Georgia and its vicinity». Philosophical Transactions of the Royal Society of London B 300: 75-106.
- Lequette, B., Verheyden, C., Jowentin, P. (1989). Olfaction in Subantarctic seabirds: Its phylogenetic and ecological significance. Condor 91: 732—135.
- Harrison, C. S. (1990) Seabirds of Hawaii, Natural History and Conservation Ithica:Cornell University Press, ISBN 0-8014-2449-6
- Grémillet, D, Chauvin, C, Wilson, R.P., Le Maho, Y. & Wanless, S. (2005). Unusual feather structure allows partial plumage wettability in diving great cormorants Phalacrocorax carbo. Journal of Avian Biology 36 (1): 57–63.
- Withers, P.C (1979). Aerodynamics and Hydrodynamics of the 'Hovering' Flight of WILSON'S Storm Petrel 80. с. 83–91. Процитовано 7 січня 2011. Проігноровано невідомий параметр
|jourbal=(довідка) - Metz, V. G., and E. A. Schreiber (2002). Great Frigatebird (Fregata minor). The Birds of North America. 681: A. Poole and F. Gill, eds. The Birds of North America, Inc. Проігноровано невідомий параметр
|city=(довідка) - Brooke, M. (2004). Albatrosses And Petrels Across The World. Oxford, UK: Oxford University Press. ISBN 0-19-850125-0.
- Gaston, Anthony J. & Jones, Ian L. (1998). The Auks. Oxford: Oxford University Press. ISBN 0-19-854032-9.
- Weimerskirch, H., Cherel, Y. (1998). Feeding ecology of short-tailed shearwaters: breeding in Tasmania and foraging in the Antarctic?. Marine Ecology Progress Series 167: 261—274.
- Prince, P.A., Huin, N., Weimerskirch, H. (1994). Diving depths of albatrosses. Antarctic Science 6 (3): 353—354.
- Ropert-Coudert Y., Grémillet D., Ryan P., Kato A., Naito Y. & Le Maho Y. (2004). Between air and water: the plunge dive of the Сарі Gannet Morus capensis. Ibis 146 (2): 281—290.
- Elliot, A. (1992). Family Pelecanidae (Pelicans) in Handbook of Birds of the World 1. Barcelona: Lynx Editions. ISBN 84-87334-10-5.
- Ainley, D.G. (1977) «Feeding methods in seabirds: а comparison of polar and tropical nesting communities in the eastern Pacific Ocean». In: Llano, G.A. (Ed.). Adaptations within Antarctic ecosystems. Smithsonian Inst. Washington D.C. P 669—685.
- Haney, J.C. & Stone, A.E. (1988). Seabird foraging tactics and water clarity: Are plunge divers really in the clear?. Marine Ecology Progress Series 49: 1–9.
- Au, D.W.K. & Pitman, R.L. (1986). Seabird interactions with Dolphins and Tuna in the Eastern Tropical Pacific. Condor 88: 304—317. Архів оригіналу за 5 листопада 2011. Процитовано 7 січня 2011.
- Schnell, G., Woods, B & Ploger B (1983). Brown Pelican foraging success and kleptoparasitism by Laughing Gulls. Auk 100: 636–644.
- Gaston, A. J., and S. B. C. Dechesne. (1996). Rhinoceros Auklet (Cerorhinca monocerata). In The Birds of North America, No. 212 (A. Poole and F. Gill, eds.). The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologists’ Union, Washington, D.C.
- Vickery, J & Brooke, M. (1994) "The Kleptoparasitic Interactions between Great Frigatebirds and Masked Boobies on Henderson Island, South Pacific " Condor 96: 331—340
- Croxall, J.P. & Prince, P.A. (1994). «Dead or alive, night or day: how do albatrosses catch squid?» Antarctic Science 6: 155—162.
- Punta, G, Herrera, G. (1995). Predation by Southern Giant Petrels Macronectes giganteus on adult Imperial Cormorants Phalacrocorax atriceps. Marine Ornithology 23: 166–167. Процитовано 7 січня 2011.
- MacArthur, R. and Wilson, E. O. (1967). The Theory of Island Biogeography, Princeton University Press (2001 reprint), ISBN 0-691-08836-5
- Плюснин Ю. М. Проблема биосоциальной эволюции: Теоретико-методологический анализ. Новосибирск. Наука. Сиб. отд-ние. 1990. ISBN 5-02-029596-5
- Robertson, C.J.R. (1993). «Survival and longevity of the Northern Royal Albatross Diomedea epomophora sanfordi at Taiaroa Head» 1937-93. Emu 93: 269—276.
- Manuwal, D. A. and A. C. Thoresen. 1993. Cassin's Auklet (Ptychoramphus aleuticus). In The Birds of North America, No. 50 (A. Poole and F. Gill, Eds.). Philadelphia: The Academy of Natural Sciences; Washington, D.C.: The American Ornithologists’ Union.
- Metz, V.G. & Schreiber, E.A. (2002) «Great Frigatebird (Fregata minor)» In The Birds of North America, No 681, (Poole, A. & Gill, F., eds) The Birds of North America Inc:Philadelphia
- Pickering, S.P.C., & Berrow, S.D., (2001) «Courtship behaviour of the Wandering Albatross Diomedea exulans at Bird Island, South Georgia» Marine Ornithology 29: 29-37
- Schreiber, E. A., C. J. Feare, B. A. Harrington, B. G. Murray, Jr., W. B. Robertson, Jr., M. J. Robertson, and G. E. Woolfenden. 2002. Sooty Tern (Sterna fuscata). In The Birds of North America, No. 665 (A. Poole and F. Gill, eds.). The Birds of North America, Inc., Philadelphia, PA.
- Seto, N. W. H., and D. O'Daniel. (1999) Bonin Petrel (Pterodroma hypoleuca). In The Birds of North America, No. 385 (A. Poole and F. Gill, eds.). The Birds of North America, Inc., Philadelphia, PA.
- Bried, J.L., Pontier, D., Jouventin, P. (2003). [www.cefe.cnrs.fr/ecomp/PDF/2003MateFidelProcAnBeh.pdf Mate fidelity in monogamous birds: а re-examination of the Procellariiformes] Перевірте схему
|url=(довідка). Animal Behaviour 65: 235—246. Процитовано 7 січня 2011. - Fisher, H.I. (1976). Some dynamics of а breeding colony of Laysan Albatrosses. Wilson Bulletin 88: 121—142.
- Rabouam, C., Thibault, J.-C., Bretagnole, V. (1998). Natal Philopatry and Close Inbreeding in Cory’s Shearwater (Calonectris diomedea). Auk 115 (2): 483—486. Архів оригіналу за 25 жовтня 2007. Процитовано 7 січня 2011.
- Moors, P.J.; Atkinson, I.A.E. (1984). Predation on seabirds by introduced animals, and factors affecting its severity.. In Status and Conservation of the World's Seabirds. Cambridge: ICBP. ISBN 0-946888-03-5.
- Keitt, B.S., Tershy, B.R. & Croll, D.A (2004). Nocturnal behavior reduces predation pressure on Black-vented Shearwaters Puffinus opisthomelas. Marine Ornithology 32 (3): 173—178. Процитовано 7 січня 2011.
- Burness, G. P., K. Lefevre, and C. T. Collins. 1999. Elegant Tern (Sterna elegans). In The Birds of North America, No. 404 (A. Poole and F. Gill, eds.). The Birds of North America, Inc., Philadelphia, PA.
- Shaffer S.A., Tremblay Y., Weimerskirch H., Scott D., Thompson D.R., Sagar P.M., Moller H., Taylor G.A., Foley D.G., Block B.A., Costa D.P. (2006). Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. Proc Natl Acad Sci. 103 (34): 12799–12802.
- Oro, D., Cam, E., Pradel, R. & Martinetz-Abrain, A. (2004). Influence of food availability on demography and local population dynamics in а long-lived seabird. Proc. R. Soc. London B 271: 387–396.
- Croxall, J, Steele, W., McInnes, S, Prince, P. (1995). Breeding Distribution of Snow Petrel Pagodroma nivea. Marine Ornithology 23: 69–99.
- Nelson, S. K. 1997. Marbled Murrelet (Brachyramphus marmoratus). In The Birds of North America, No. 276 (A. Poole and F. Gill, eds.). The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologists’ Union, Washington, D.C.
- Winkler, D. W. 1996. California Gull (Larus californicus). In The Birds of North America, No. 259 (A. Poole and F. Gill, eds.). The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologists' Union, Washington, D.C.
- Harris, M. & Wanless, S. year = 1996. Differential responses of Guillemot Uria aalge and Shag Phalacrocorax aristotelis to а late winter wreck. Bird Study 43 (2): 220—230.
- Collis, K., Adamany, S. (Columbia River Inter-Tribal Fish Commission) Roby, D.D., Craig, D.P., Lyons, D.E. (Oregon Cooperative Fish and Wildlife Research Unit) (2000). Avian Predation on Juvenile Salmonids in the Lower Columbia River. 1998 Annual Report to Bonneville Power Administration, Portland, OR. Архів оригіналу за 22 червня 2013. Процитовано 11 червня 2008.
- Oro, D., Ruiz, X., Pedrocchi, V. & Gonzalez-Solis, J. (1997) «Diet and adult time budgets of Audouin's Gull Larus audouinii in response to changes in commercial fisheries» Ibis 139: 631—637
- Thompson, P.M., (2004) «Identifying drivers of change; did fisheries play а role in the spread of North Atlantic fulmars?» in Management of marine ecosystems: monitoring change in upper trophic levels. Cambridge: Cambridge University Press Архівовано 23 серпня 2006 у Wayback Machine.
- Save the Albatross: The Problem. BirdLife International/RSPB. 2005. Архів оригіналу за 22 червня 2013. Процитовано 17 березня 2006.
- Brothers NP. 1991. «Albatross mortality and associated bait loss in the Japanese longline fishery in the southern ocean.» Biological Conservation 55: 255—268.
- С. И. Корнев. «Проблемы сохранения морских млекопитающих и птиц при рыбном промысле в российских водах северной части Тихого океана» онлайн
- Simeone, A. & Navarro, X. (2002) «Human exploitation of seabirds in coastal southern Chile during the mid-Holocene» Rev. chil. hist. nat 75(2): 423—431
- Anderson, A. (1996) «Origins of Procellariidae Hunting in the Southwest Pacific» International Journal of Osteoarcheology 6: 403—410
- White, Peter; (1995), The Farallon Islands, Sentinels of the Golden Gate, Scottwall Associates:San Francisco, ISBN 0-942087-10-0
- Burnham, W., Burnham, K.K., Cade, T.J., (2005) «Past and present assessments of bird life in Uummannaq District, West Greenland» Dansk Orn. Foren. Tidsskr. 99: 196—208
- Carlile, N., Proiddel, D., Zino, F., Natividad, C. & Wingate, D.B. (2003) «A review of four successful recovery programmes for threatened sub-tropical petrels» Marine Ornithology 31: 185—192
- Fry, D. & Toone, C. (1981) «DDT-induced feminization of gull embryos» Science 213(4510): 922—924
- Dunnet, G., Crisp, D., Conan, G., Bourne, W. (1982) «Oil Pollution and Seabird Populations [and Discussion]» Philosophical Transactions of the Royal Society of London. B 297(1087): 413—427
- USFWS Pelican Island National Wildlife Refuge History of Pelican Island Retrieved September 2 2006
- Williams, J.C., Byrd G.V.& Konyukhov, N.B. (2003) «Whiskered Auklets Aethia pygmaea, foxes, humans and how to right а wrong.» Marine Ornithology 31: 175—180
- BirdLife International (2005) Stamps celebrate seabird return. Retrived August 12 2006
- Food and Agriculture Organisation (1999) "The incidental catch of seabirds by longline fisheries: worldwide review and technical guidelines for mitigation. FAO Fisheries Circular No.937. Food and Agriculture Organization of the United Nations, Rome.
- Australian Antarctic Division Agreement on the Conservation of Albatrosses and Petrels Site Retrieved September 2 2006
- Carboneras, C. (1992) «Family Diomedeidae (Albatrosses)» in Handbook of Birds of the World Vol 1. Barcelona:Lynx Edicions, ISBN 84-87334-10-5
Ресурси Інтернету
- BirdLife International: Кампанія по збереженню альбатросів (англ.)
- Marine Ornithology, журнал по вивченню і охороні морських птахів (англ.)
- Seabirds, Foragefish, and Marine Ecosystems Research. Архів оригіналу за 22 червня 2013. Процитовано 7 січня 2011. Текст «USGS» проігноровано (довідка) (англ.)
- Alaska Seabird Information Series. Архів оригіналу за 22 червня 2013. Процитовано 14 червня 2008. (англ.)
- С. И. Корнев. Проблемы сохранения морских млекопитающих и птиц при рыбном промысле.... ФПС РФ. Архів оригіналу за 22 червня 2013. Процитовано 11 червня 2008. (рос.)
- Ю. Б. Артюхин, А. В. Винников, Д. А. Терентьев. Морские птицы и донное ярусное рыболовство. Архів оригіналу за 12 липня 2007. Процитовано 14 червня 2008. (рос.)
- В.И.Придатко. Морские колониальные птицы островов Врангеля и Геральд. Архів оригіналу за 1 червня 2013. Процитовано 20 березня 2010. (рос.)
|
|
Ця стаття належить до добрих статей української Вікіпедії. |